|
ヒメアカタテハやクロヒカゲの占有行動は交尾の為ではない
(序でに、蝶界への疑問)
遠藤英實 作
目 次
0 はじめに
1 占有行動の概略
2 2018年の個体数
3 占有域(A樹、B樹)説明
4 占有行動の態様
5 ヒメアカタテハの発生回数(準備中)
6 終わりに
0章 はじめに
ヒメアカタテハの観察は今年(2018年)で3年目を終えた。
その結果、私は
ヒメアカタテハの占有行動は交尾の為ではない
と考えたい。
調査域は1ヶ所のみであったが、
3年間一度もそれらしい行動を目撃出来なかったからである。
それらしい行動とは
待ち構えていた雄が、傍にやってきた雌に近づき交尾をする。
或いはそれらしい行動をとる
ことである
私の調査域は1ヶ所であったが、他者の観察情報(真面目な自己の観察結果)でも
然りである。
クロヒカゲの観察は、今年は行わなかったが、
クロヒカゲの占有行動も、ヒメアカタテハと殆ど同様である。
クロヒカゲの観察域は東京1ヶ所、盛岡市2か所であるが、
どちらもそれらしい行動を目撃できなかった。
盛岡市の場合は、占有域すら発見出来なかったのである。
某プロの“論説”では、
“クロヒカゲの占有行動―>交尾”らしいが、
このプロがそもそも交尾を目撃していないのは、
現場で観察してきた人間(例えば私)にはすぐ分かる。
現場で本当に観察したのかどうかさえ疑わしい。
然らば、このプロがどうしてかような“論説”をものしたかというに、
舶来の“学術用語”、レック型、非レック型という言葉を
を使いたかったからではなかろうか?
日本で観察出来るのなら、日本で観察しろ!と云いたい。
自己の“論文”の権威付けのために
舶来の学術用語や舶来の文献利用は止めるべきである。
日本人の納める税金で“研究”させて貰っているのであろうから!!
1 占有行動の概略
ヒメアカタテハの占有行動を調べて3年経った。
そろそろ纏めてみたい。
観察域はこれまでと同じである。
結局、以下の2か所(A樹、B樹)のみが占有域であった。
(A樹、B樹)については後段で図示する。
以下概略。
① この2か所のみで、集結開始時期から姿を消す時期まで、
ほぼコンスタントに占有行動を目撃することが出来る。
つまり、A樹、B樹それぞれの根方で静止していて、
他蝶が近づくと、直ちに追い払うのである。
ほかの各所でも争いは、目撃は出来るのだが、
・一過性である(例えば、行きに目撃して、帰りには姿を消している)
・スケールが小さく、バトルではなく「小競り合い程度」である
という点で、占有行動とは云い難い。
② ヒメアカタテハは、この観察域内で移動している。以下、例。
広い観察域内の下流で10数匹が吸蜜していた。
少しずつ数が減り、4~5日後には0匹となっていた。
(その代わりと云っては何だが)
上流で10数匹が吸蜜していた。
この域内で移動したのは確かであろう。
(ほぼ数が合っているから)
この域内で死ぬのか?
それとも、更に他所へ移動するのか?
分からない。(多分殆どは前者)
面白いのは、広い観察域内で移動していても、
A樹、B樹に最後まで(12月迄)目撃される
ことである。つまり、
A樹、B樹の根方に惹かれる
のだと思う。
根方にしがみついてひたすら(寒い12月迄)雌を待っているわけではなかろう。
③ 占有行動は交尾行動と結びついてはいない。
私がこの3年間で目撃した交尾は、
A樹-B樹間のブロック道(約100m)の 両側のみであった。
A樹、B樹の根方で待ち構えている雄に雌が近づいていって、
交尾する光景など一度も目撃したことはない。
レック型なる“専門用語”は、クロヒカゲの場合と同様に、
観察なんぞしていない学者の絵空事であると思う。
④ 同種、異種間の争いは、この2か所以外でも頻繁に見られるのであるが、
それは小競り合い程度である。人間であれ、蝶であれ、近づき過ぎたら鬱陶しい。
また、占有域を広く設定するのも良くないと思う。
何故なら、広い範囲では、小競り合いはあちこちで起こっているから。
⑤ 異種間の争いは、瞬時に収まる。
(対アゲハチョウ、アカボシゴマダラ、キタテハ、モンシロチョウ、モンキチョウ
キチョウ、ヤマトシジミ)
但し、イチモンジセセリは例外である。
ヒメアカタテハのバトルに、イチモンジセセリが介入してくる雰囲気なのである。
大きさは全く違うのだから、雄が雌と間違えての筈はなかろう。
“飛翔速度が似ているから惹き付けられる”と云いたいところである。
(イチモンジセセリとの場合は、広い範囲でバトルが繰り広げられるわけではないのだが、
私でも撮れる範囲には広がる。)
⑥ 同種間の争いは様相を異にする。
後段で図示するが、スケールが大きくなり、
多摩川の向こう岸まで繰り広げられることがある。
このバトルは多分雄同士であると思う。
大空を駆け巡るスケールの大きいバトルは、A樹、B樹絡みに限られる。
⑦ 雌雄の争いも見かけるが、このようにスケールの大きいバトルではない。
地面近くを雌が逃げ回り、雄が(場合によっては執拗に)追いかけ回す程度である。
つまり、普通の交尾拒否行動である。
なお交尾拒否行動は、A樹-B樹間のブロック道だけではなく、
周辺の草むらでも目撃出来る。
また、A樹-B樹間のブロック道での交尾拒否行動と思っていたら、
結局B樹で交尾したケースが1例だけあった。
これは、雌にとっては、“力尽きての無念の結果”ではなかろうか。
他種でも、このような結果をしばしば目撃している。
⑧ A樹、B樹での占有行動中に個体は入れ替わるのだろうか?
新しい個体と古びた個体とで簡単に確認出来る場合がある。
例えば、
・前半は新しい個体、後半は古びた個体 で入れ替わっていることが確認出来た。
・更に、新しい個体に変わることがあった。
日付が変わると、個体が変わっている場合が多いようだ。
マーキングすれば正確に分かりそうに思えるけれど
多分失敗に終わるだろう。痛めつけられた蝶は行方を晦ますと思うのである。
つまり、マーキング作業は、結果に大きな改変をもたらすと思う。
また、中筋 房夫氏はイチモンジセセリの論文で、
「5年間で1万匹にマーキングして回収は4匹」
と述べている。
つまり、統計処理上は全く無意味な作業なのである。
(飛行距離の調査としては面白いが。)
⑨ A樹、B樹は睡眠と関係があるか?
一度だけ、A樹、B樹でそれぞれ睡眠に入った個体があった。
大抵は、日暮れ時にどこかへ飛び去っていった。
このことも、その場所が交尾の為とは考えづらい理由である。
そこが交尾に好都合の場所なら、そのままそこで睡眠に入れば良いではないか。
(次の日、直ぐ占有出来るから。)
ところが夕方姿を消し、次の日は入れ替わっているように思える。
そして次の日の個体も、やはり強いのである。
⑩ 固執するエリアは極狭い範囲である。
10mも離れていない隣の樹の個体には全く無関心である。
交尾の為の占有なら、追っ払っても良さそうなのに!
⑪ 最後の大きな疑問は、
《その占有域が毎年同じ場所であること》
である。
その場所を探す手掛かりは何なのか?
後述するが、ミツバチの交尾場所も毎年決まっている。
ミツバチが集合場所を探す手掛かりは今のところ、
「目立つ目印や特徴的な地形、つまり視覚的なもの」
とされている。
それならば、ヒメアカタテハの占有域を示す手がかりも
「視覚的なもの」
と考えられるのではなかろうか。
(目的は、ミツバチと違って交尾行動ではないが。)
つまり(私の観察域の場合)、
・お気に入りのブロック道
・両側の草むら(ヨモギ、コセンダングサ<-この蜜を好む)
・両端の樹木(A樹、B樹)
・この光景を捉えて、象徴的な存在である両端の樹木に集まる
と考えるわけである。
(なお、ブロック道の中間に他の樹木もあるが、これには全く関心を示さない。)
2 2018年の個体数
異常気象(気象庁発表)のせいか、個体数は過去の半分以下である。
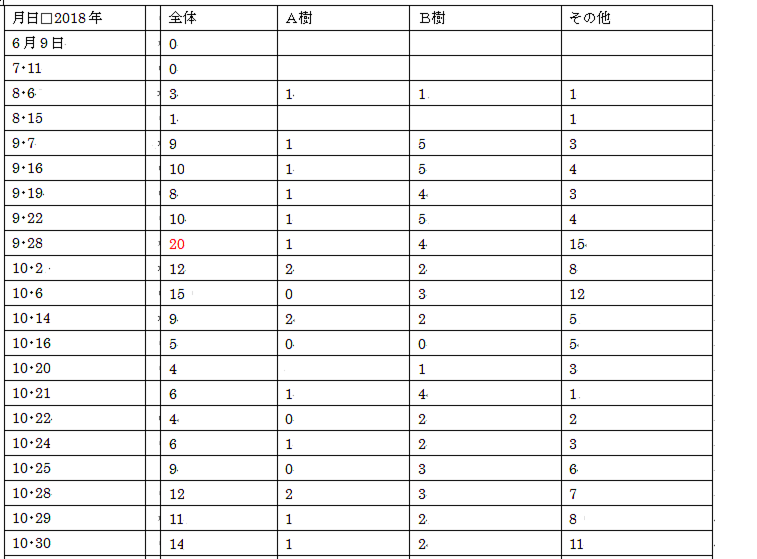
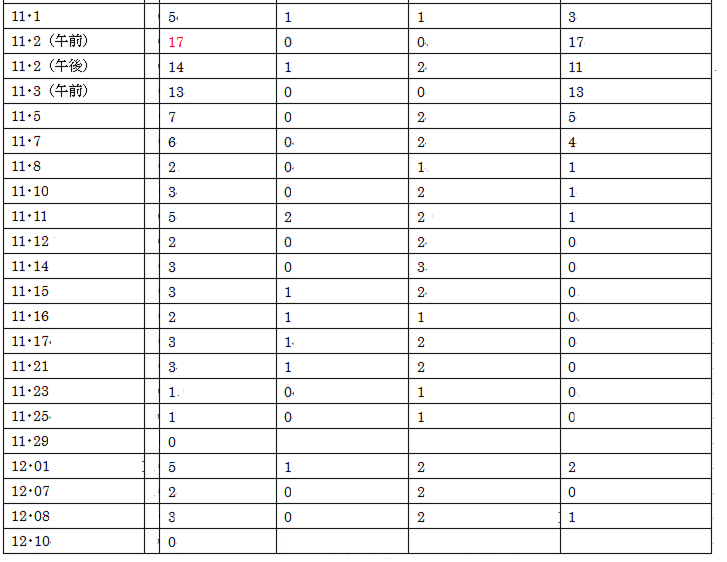 (B樹については、廻りの樹、及び周辺のブロック土手から侵入してくる個体も含める。)
感想①
気象庁発表の狛江市の気温は、12・10以降は気温10℃以下、但し12・17以降は12℃以上である。
12・19、観察地で実際に計ってみた。14℃~22℃である。
当然のことではあるが、「晴れ」であっても、
天候の状態(風速、雲がかかっているか、etc)、計る場所(日向は日陰か、etc)
によって全く違ってくるのである。
だから私は、論文などに実験室での気温や照度が麗々しく大げさに登場すると違和感を覚える。
蝶は実験室内の動物ではない! 野外の動物なのだ!
感想②
個体数を数える時間帯は概ね午後である。
11月2日の午前、午後の記録を見ても分かるように、占有行動は午後から始まる。
午前中は周辺で遊んでいるようだ。
午後になって一部が、或いはタイムシェアで相当数が、占有行動をとる。
A樹では、他個体が周囲の草むらから襲ってくる。
B樹では、他個体が廻りの狭い範囲(隣の樹、奥の樹、近くのブロック道、etc)から襲ってくる。
感想③
この観察域への登場から姿を消す迄、A樹、B樹で占有行動を目撃出来たことに驚かされる。
B樹から姿を消すこと=この観察域から姿を消すこと
なのだ。
他の行動と合わせて考えても、ひたすら雌を求めてというのは考えづらい。
この場所が気に入っていると考えるのが自然である。
アカタテハやルリタテハも毎年ほぼ同じポイント(よりは広い)に1,2匹が現れるが
1日(或いは2~3日)で姿を消す。
偶々2匹だと猛烈なバトルを展開する。
しかしこれを、雌を巡る占有行動とは云わないだろう。
ヒメアカタテハの場合は、期間が長く(最初から最後迄)、数多くが参加するのである。
感想④
ヒメアカタテハの場合、各個体の占有期間はあまり長くないようだ。
その日は死守しても、次の日は入れ替わっている場合がある。
また、その日のうちに入れ替わっている場合もある。
(個体の破損状況で分かる。)
自から放棄するのか? 他者から追い払われるのか?
感想⑤
占有域との関係は、クロヒカゲと良く似ている。
クロヒカゲも、観察域への登場から退場までほぼ全期間、
占有域で目撃出来る。
そして、他の場所では姿を消しても、占有域では目撃出来る。
占有域では、他個体が襲ってこない限り、
ひたすら至福の(?)時を過ごす。
やがて、日没が迫ると、自らも姿を消す。
“ひたすら交尾を求めて”だのレック型だの、プロの作り話だ!
感想⑥
今年は、異常気象(気象庁発表)だったことは、私も実感した。
この地で観察を楽しんでいたヒメアカタテハ、コムラサキ、ヒメウラナミジャノメが
異常に少なかったのである。
ヒメウラナミジャノメは、他の観察地でもまさに気息奄々の状態であった。
ヒメアカタテハの個体数も表で示したように過去の半分以下、
コムラサキはもともと樹上の蝶であるから個体数は把握し難いのだが、
活発に飛び回っているという印象はまるでなかった。
その代わり(と云っては何だが)、
イチモンジセセリとウラナミシジミは、今年は異常に多かった。
特に同じ調査域のウラナミシジミは、ヒメウラナミジャノメの過去のカウント時から類推して
最大500~,600匹/日以上いたと思う。例年の数倍、これは異常である。
イチモンジセセリは都心だけでも、交尾を4回目撃している。
どちらも南方系の蝶と云われているから、暑さに耐えかねて北上個体が増加したのか?
(<―インチキ推理)
来年はどうなるのだろう?
蝶は意外にしぶといから、また元に戻るような気がするが、
来年を楽しみにしている。
3 占有域(A樹、B樹)説明
前述のように、河川敷の一部にコンクリートブロックの土手がある。
その一部(約100m)が、ヒメアカタテハの特に好む場所であり、
その両端の樹A、Bの根方がそれぞれ占有域となる。
(樹そのものが占有域ではない。)
その根方の位置、面積は、人の気配、気温によって多少変化する。
(この場所は、人の遊び場でもある。)
(B樹については、廻りの樹、及び周辺のブロック土手から侵入してくる個体も含める。)
感想①
気象庁発表の狛江市の気温は、12・10以降は気温10℃以下、但し12・17以降は12℃以上である。
12・19、観察地で実際に計ってみた。14℃~22℃である。
当然のことではあるが、「晴れ」であっても、
天候の状態(風速、雲がかかっているか、etc)、計る場所(日向は日陰か、etc)
によって全く違ってくるのである。
だから私は、論文などに実験室での気温や照度が麗々しく大げさに登場すると違和感を覚える。
蝶は実験室内の動物ではない! 野外の動物なのだ!
感想②
個体数を数える時間帯は概ね午後である。
11月2日の午前、午後の記録を見ても分かるように、占有行動は午後から始まる。
午前中は周辺で遊んでいるようだ。
午後になって一部が、或いはタイムシェアで相当数が、占有行動をとる。
A樹では、他個体が周囲の草むらから襲ってくる。
B樹では、他個体が廻りの狭い範囲(隣の樹、奥の樹、近くのブロック道、etc)から襲ってくる。
感想③
この観察域への登場から姿を消す迄、A樹、B樹で占有行動を目撃出来たことに驚かされる。
B樹から姿を消すこと=この観察域から姿を消すこと
なのだ。
他の行動と合わせて考えても、ひたすら雌を求めてというのは考えづらい。
この場所が気に入っていると考えるのが自然である。
アカタテハやルリタテハも毎年ほぼ同じポイント(よりは広い)に1,2匹が現れるが
1日(或いは2~3日)で姿を消す。
偶々2匹だと猛烈なバトルを展開する。
しかしこれを、雌を巡る占有行動とは云わないだろう。
ヒメアカタテハの場合は、期間が長く(最初から最後迄)、数多くが参加するのである。
感想④
ヒメアカタテハの場合、各個体の占有期間はあまり長くないようだ。
その日は死守しても、次の日は入れ替わっている場合がある。
また、その日のうちに入れ替わっている場合もある。
(個体の破損状況で分かる。)
自から放棄するのか? 他者から追い払われるのか?
感想⑤
占有域との関係は、クロヒカゲと良く似ている。
クロヒカゲも、観察域への登場から退場までほぼ全期間、
占有域で目撃出来る。
そして、他の場所では姿を消しても、占有域では目撃出来る。
占有域では、他個体が襲ってこない限り、
ひたすら至福の(?)時を過ごす。
やがて、日没が迫ると、自らも姿を消す。
“ひたすら交尾を求めて”だのレック型だの、プロの作り話だ!
感想⑥
今年は、異常気象(気象庁発表)だったことは、私も実感した。
この地で観察を楽しんでいたヒメアカタテハ、コムラサキ、ヒメウラナミジャノメが
異常に少なかったのである。
ヒメウラナミジャノメは、他の観察地でもまさに気息奄々の状態であった。
ヒメアカタテハの個体数も表で示したように過去の半分以下、
コムラサキはもともと樹上の蝶であるから個体数は把握し難いのだが、
活発に飛び回っているという印象はまるでなかった。
その代わり(と云っては何だが)、
イチモンジセセリとウラナミシジミは、今年は異常に多かった。
特に同じ調査域のウラナミシジミは、ヒメウラナミジャノメの過去のカウント時から類推して
最大500~,600匹/日以上いたと思う。例年の数倍、これは異常である。
イチモンジセセリは都心だけでも、交尾を4回目撃している。
どちらも南方系の蝶と云われているから、暑さに耐えかねて北上個体が増加したのか?
(<―インチキ推理)
来年はどうなるのだろう?
蝶は意外にしぶといから、また元に戻るような気がするが、
来年を楽しみにしている。
3 占有域(A樹、B樹)説明
前述のように、河川敷の一部にコンクリートブロックの土手がある。
その一部(約100m)が、ヒメアカタテハの特に好む場所であり、
その両端の樹A、Bの根方がそれぞれ占有域となる。
(樹そのものが占有域ではない。)
その根方の位置、面積は、人の気配、気温によって多少変化する。
(この場所は、人の遊び場でもある。)
 A樹(上部から) 横から
黒い枠の樹がA樹であり、その周囲をヒメアカタテハは好む。
ヒメアカタテハは通常上部の土手を縄張りとするが、秋になると暖かい壁側に止まるようになる。
A樹(上部から) 横から
黒い枠の樹がA樹であり、その周囲をヒメアカタテハは好む。
ヒメアカタテハは通常上部の土手を縄張りとするが、秋になると暖かい壁側に止まるようになる。
 B樹(赤) 手前はA樹 A樹、B樹間は100m位 中間にある樹
赤い枠が、次なる縄張りのB樹である。
ヒメアカタテハは青枠の樹の周りにも止まっているが
青枠の樹は縄張りとは云えない。
赤枠側は関心を示さないのである。
このことも、
占有行動は雌を求める行動ではない
と私が考える理由の一つ(強い理由の一つ)である。
A樹、B樹間は10mもない。
探雌行動なら、A樹、B樹同じ位置のようなものだから
果敢に追い立てる筈ではないか。
逆に、青枠側は時々赤枠側に襲い掛かって来て撃退される。
(マーキングで確かめたわけではないから、
“撃退される”というのは“多分”である。
マーキング作業をしたら、多分その個体はいなくなってしまうだろう。
A樹、B樹間は約100mである。
両側に食草ヨモギや好物のコセンダングサがびっしりと生えている。
これまで観察した交尾行動は全てこの範囲である。
この区間は、云わば共有区間であり縄張りとは云えない。
確かに、この共有区間でも諍いはあるが、その諍いは一過性であり、しかも瞬間的に収まる。
また、この共有区間の中間の位置に小樹があるが、この小樹には全く関心を示さない。
A樹、B樹に関しては、とにかく縄張りを守るという意思が明確なのであるが、後述。
(この樹の種類は私には分からない。地元の人によれば種が飛んできて自然に生えたらしい。
いずれにしろ、種類は関係なかろう。やはり、「位置」が重要であると思う。)
4 占有行動の態様
① A樹占有
B樹(赤) 手前はA樹 A樹、B樹間は100m位 中間にある樹
赤い枠が、次なる縄張りのB樹である。
ヒメアカタテハは青枠の樹の周りにも止まっているが
青枠の樹は縄張りとは云えない。
赤枠側は関心を示さないのである。
このことも、
占有行動は雌を求める行動ではない
と私が考える理由の一つ(強い理由の一つ)である。
A樹、B樹間は10mもない。
探雌行動なら、A樹、B樹同じ位置のようなものだから
果敢に追い立てる筈ではないか。
逆に、青枠側は時々赤枠側に襲い掛かって来て撃退される。
(マーキングで確かめたわけではないから、
“撃退される”というのは“多分”である。
マーキング作業をしたら、多分その個体はいなくなってしまうだろう。
A樹、B樹間は約100mである。
両側に食草ヨモギや好物のコセンダングサがびっしりと生えている。
これまで観察した交尾行動は全てこの範囲である。
この区間は、云わば共有区間であり縄張りとは云えない。
確かに、この共有区間でも諍いはあるが、その諍いは一過性であり、しかも瞬間的に収まる。
また、この共有区間の中間の位置に小樹があるが、この小樹には全く関心を示さない。
A樹、B樹に関しては、とにかく縄張りを守るという意思が明確なのであるが、後述。
(この樹の種類は私には分からない。地元の人によれば種が飛んできて自然に生えたらしい。
いずれにしろ、種類は関係なかろう。やはり、「位置」が重要であると思う。)
4 占有行動の態様
① A樹占有
 A樹 ブロック側 路上の占有個体
A樹 ブロック側 路上の占有個体
 寒くなってくると壁に止まる 左10・21 右11.21
A樹では、寒くなってくると、西日の当たる壁に止まり出すのがおかしい。
ハチ、トンボ、他の蝶も止まり出す。
② B樹占有
寒くなってくると壁に止まる 左10・21 右11.21
A樹では、寒くなってくると、西日の当たる壁に止まり出すのがおかしい。
ハチ、トンボ、他の蝶も止まり出す。
② B樹占有
 B樹を占有する 占有個体
B樹には、隣の青い樹、手前のブロック道、奥の繁みからも集まってくる(襲ってくる)。
多分、それらは占有域ではない。B樹の蝶は関心を示さないのである。
B樹を占有する 占有個体
B樹には、隣の青い樹、手前のブロック道、奥の繁みからも集まってくる(襲ってくる)。
多分、それらは占有域ではない。B樹の蝶は関心を示さないのである。
 B樹と隣の青枠の樹の間の草むらである。
夕方になると2匹が仲良く止まっている。そのうちどちらも姿を消す。
③ 同種間の争い(雄と雄)
B樹と隣の青枠の樹の間の草むらである。
夕方になると2匹が仲良く止まっている。そのうちどちらも姿を消す。
③ 同種間の争い(雄と雄)
 A樹を巡るバトル
A樹を巡るバトル
 B樹を巡るバトル
かくの如く、壮大なバトルが繰り広げられる。多摩川の対岸にまで及ぶ時もある。
多分雄同士であると思う。このようなスケールの大きいバトルは、他種でも目撃した記憶はない。
B樹を巡るバトル
かくの如く、壮大なバトルが繰り広げられる。多摩川の対岸にまで及ぶ時もある。
多分雄同士であると思う。このようなスケールの大きいバトルは、他種でも目撃した記憶はない。
 対イチモンジセセリ 2対4 2対1
時にはイチモンジセセリも参戦する。
概ね、ヒメアカタテハが彼方に飛び去って闘いは終わる。
④ 同種間の争い(雄と雌)
雌雄は、前項のような闘いをしないと思う。
通常の交尾拒否ではなかろうか。
それならば、ブロック間でもそれ以外の草むらでもしばしば目撃している。以下、例。
対イチモンジセセリ 2対4 2対1
時にはイチモンジセセリも参戦する。
概ね、ヒメアカタテハが彼方に飛び去って闘いは終わる。
④ 同種間の争い(雄と雌)
雌雄は、前項のような闘いをしないと思う。
通常の交尾拒否ではなかろうか。
それならば、ブロック間でもそれ以外の草むらでもしばしば目撃している。以下、例。
 15時29分
15時29分
 撮れず 15時35分
交尾拒否を目撃した。9月28日15時29分。
雄は執拗に迫り、雌は断固として拒否する。
そのうち雌は草むらを逃げ出した。雄は追走する。
飛び回っているが撮れない。結局、15時35分草原の彼方に共に姿を消した。
この間6分、見物していると結構長い。
これが雌雄の諍いではなかろうか。バトルとは言い難い。
⑤ 異種間の争い
撮れず 15時35分
交尾拒否を目撃した。9月28日15時29分。
雄は執拗に迫り、雌は断固として拒否する。
そのうち雌は草むらを逃げ出した。雄は追走する。
飛び回っているが撮れない。結局、15時35分草原の彼方に共に姿を消した。
この間6分、見物していると結構長い。
これが雌雄の諍いではなかろうか。バトルとは言い難い。
⑤ 異種間の争い
 対モンキチョウ
異種間の争いはしばしば見かけるのだが、瞬間的で私にはなかなか撮れない。
かろうじて、対モンキチョウは撮れた。
どれも、ヒメアカタテハが圧倒する。(つまり異種側が直ちに飛び去る。)
⑥ 占有域での個体の移り変わり
占有域内で、占有個体は変わっているか?
対モンキチョウ
異種間の争いはしばしば見かけるのだが、瞬間的で私にはなかなか撮れない。
かろうじて、対モンキチョウは撮れた。
どれも、ヒメアカタテハが圧倒する。(つまり異種側が直ちに飛び去る。)
⑥ 占有域での個体の移り変わり
占有域内で、占有個体は変わっているか?
 B樹 10月22日 多少ボロ B樹 10月24日 美麗
日が違えば個体も変わっているのが分かる。(常に変わるわけではないが。)
B樹 10月22日 多少ボロ B樹 10月24日 美麗
日が違えば個体も変わっているのが分かる。(常に変わるわけではないが。)
 B樹(赤い枠)11月05日 隣の樹(青い枠)11月05日
左の占有個体(美麗)を右の個体(ボロ)が襲った。直ぐ撃退された。
時々、ボロが襲ってくるがやはり撃退される。美麗とボロの場合は分かり易い。
B樹(赤い枠)11月05日 隣の樹(青い枠)11月05日
左の占有個体(美麗)を右の個体(ボロ)が襲った。直ぐ撃退された。
時々、ボロが襲ってくるがやはり撃退される。美麗とボロの場合は分かり易い。
 B樹 やや美麗 B樹 別個体 B樹 元の個体
別の日、B樹を離れて戻ってきたら占有個体が変わっていた。(やや美麗からボロへ)
そのまま見ていると、すぐ元の個体が戻って来てボロの個体を追い払った。
元の個体は、奥の場所に吸蜜に行っていたように思えた。
B樹 やや美麗 B樹 別個体 B樹 元の個体
別の日、B樹を離れて戻ってきたら占有個体が変わっていた。(やや美麗からボロへ)
そのまま見ていると、すぐ元の個体が戻って来てボロの個体を追い払った。
元の個体は、奥の場所に吸蜜に行っていたように思えた。
 B樹 美麗 ボロ 美麗
別の日、いつの間にか変わっていたが、そのタイミングは分からなかった。
B樹 美麗 ボロ 美麗
別の日、いつの間にか変わっていたが、そのタイミングは分からなかった。
 A樹壁 破損 14時59分 完品15時34分
別の日、いつの間にか入れ替わっていた。
⑦ 単なる休息
A樹壁 破損 14時59分 完品15時34分
別の日、いつの間にか入れ替わっていた。
⑦ 単なる休息
 占有域以外では、こういう光景も良く見かける。
路上は好みの場所である。
但し遺伝子に組み込まれた占有域ではない。
だから派手な喧嘩はしない?
⑧ 総括
かくの如く、集結し始めてからやがて姿を消す迄、いずれかの個体が占有域に陣取っている。
占有個体は、そのポイントを死守しようとするが、頻繁に入れ替わっているようでもある。
その間、占有域で交尾行動を目撃したことはない。
どうみても、占有行動は雌確保の為ではない。
“その場所”が気に入っているからと考えた方が良さそうだ。
“その場所”は極狭い範囲である。
ちょっと離れると見向きもしない。
成蝶の行動は、採餌、交尾、産卵、休息、睡眠から成るが、
占有行動は、気に入った場所での休息と思う。
そこを確保するためには、闘いも辞さず というのが可笑しい。
5 ヒメアカタテハの発生回数(別サイトへ移行)
6 終わりに
蝶界への疑問を脈絡もなく考えている。
例えば、
・蝶道 ・占有行動 ・ヒルトッピング ・交尾飛翔形式
・野外での観察と実験室での“研究” ・標本の所有者と“研究者”
・1化と遺伝的1化とダラダラ1化
など。
今回は、ヒメアカタテハの占有行動と交尾との関係について述べた。
他に考えていることを述べてみる。
① アオバセセリの占有行動
アオバセセリについて述べた論文を読んだ。
それによれば、
観察期間中、縄張りでの交尾を1回目撃した。
他にもいろいろの状況証拠を揃えた。
よって、
占有行動―>交尾
と結論づけた
ということらしいが、私にはさっぱり分からない。
・0回目撃なら氏は自説を撤回するのだろうか?
然らば、0回と1回には天と地程の違いがあるが
そんな違いはあるのだろうか?
・例えば、
「アオバセセリとスミナガシは同じ位の大きさだから
雌と誤認して執拗に追いかける」
なる観察がある。
どうして、これらが状況証拠になるのだろうか?
アオバセセリの占有行動も交尾とは関係がないだろう。
「山頂に惹かれて集まってくる」のだと思う。
② 蜂の話題
ミツバチの生態について書かれている著書、ウェブサイトは多い。
配偶行動について取りあえず私が紹介する。(不正確かも知れない)
*************************************
・春、雄バチ群が羽化する。1日数回、巣を出入りする。
交尾場所を探索、確認しているらしい。
・そのうち、女王バチ候補が羽化する。
雄バチは、同じ巣の女王バチ候補には全く関心を示さない。
つまり、近親交配を忌避しているのである。
・各巣から出陣した雄バチは、特定の交尾場所に集結している。
女王バチ候補が性フェロモンを振りまきながら現れると、
目敏く見つけた雄群が黒い塊となって女王バチ候補を追いかけ
運の良い雄が交尾をする。
・雄バチは1回の交尾で死ぬが、女王バチ候補は、7~8回交尾をするらしい。
(ということは、子どもの働きバチの父親は7~8匹いることになる)
・問題は交尾場所である。白昼、黒い塊が唸りを上げて飛び回っている。
高さ:10~40m 直径:50~200m
極普通に飛んでいるミツバチなのだから、
素人でも交尾場所は発見出来そうだが、これが相当の難問らしい。
各国の発見例が紹介されていて、日本では未発見となっていた。(或る著書では)
その後、日本でもハイテク機器を駆使して発見されたようだ。
・もう一つの難問は、《その交尾場所が毎年同じ場所であること》である。
ハチが集合場所を探す手掛かりは今のところ、
目立つ目印や特徴的な地形、つまり視覚的なもの とされている。
・トウヨウミツバチ(亜種ニホンミツバチ)、スズメバチについても
いろいろな研究が報告さている。
***************************************
読んでいて、私のような素人でも興奮するのである。
目下、二つの点が気になっている。
一つ目は、ハチは今や家畜であるということである。
養蜂家としては、
「働きの良い巣をつくりたい。
女王バチが勝手に外に出かけていって、
体力はあるが怠け者のオスと勝手に交尾してもらっては困る。」
だから、近親交配は避けるというハチが守ってきたルールは
養蜂家によって破られているようなのである。
それがハチの反乱、即ち
「もう、かんべんしてくれ!」ということで、
働きバチが巣に帰ってこない--->巣の崩壊
をもたらしているのではないだろうか?
(実際はどうなっているか分からないけれど)
二つ目は、以下の指摘である。
「ハチが集合場所を探す手掛かりは今のところ、
目立つ目印や特徴的な地形、つまり視覚的なものである。」
つまり、「このような光景へ惹かれること」が遺伝子に組み込まれていると云うことだ。
毎年産まれてきた雄や女王バチ候補が同じ場所に集まってくるのだから、
こう考えざるを得ないということだろう。
こういう論法は、蝶の行動(一部の種の一部の個体の行動)にも当てはまるのではなかろうか。
例えば、山登り(ヒルトッピング)、占有行動。
(但し蝶の場合は、このことと配偶行動とは関係がないと私が思う所以は後段で。)
③ 蝶の話題
然らば、蝶の場合はどうであろうか?
ヒルトッピング、レック型、非レック型なる“専門用語”がある。
これらに関する記述は、いろいろな著書やウェブサイトで見られるが、
例えば、https://www.pteron-world.com/topics/ecology/mating.html
アメリカの学者シールズは、
「山に集まって来る雌は交尾していない個体で、山頂で交尾して山を下る。」ことを“証明した”らしい。
(因みに、その後多用されている「レック」というのは
蝶が交尾のために集まってくる場所をさすらしい。シールズが命名したのかな?)
アメリカ人がアメリカの山でどういう調査をしたのか知らないけれど、
そのような学説があるのなら、日本人は日本で確かめてみれば良いではないか。
私は子どもの頃から、いろいろな山に登ったけれど、
交尾が活発に行われている光景などまるで思い出せない。
多数の蝶が集まって来る極せまい山頂にも何度も登ったけれど、
そもそも交尾そのものが全く思い出せないのである。
(殆ど雄だけだったような気がする。)
往時は交尾行動に特別に関心があったわけではないが、
この十数年は、岩手県の山、東京周辺の山を調べてみた。
シールズのように「入り鉄砲に出女を調べる関所の役人」のような熱心さで
調べたわけではないが、それでもそのような事例は全く目撃出来なかった。
ウェブサイトを見ても、
「集合場所では雌は目撃できなかった、交尾は目撃出来なかった」
という報告ばかりである。
少なくとも、《蝶の「占有場所」はミツバチの「交尾空間」と同じ》なる報告は全くないのである。
私は、シールズの“証明”は、アメリカにあっても“false”であると思っている。
頂上でマーキングした雌がたまたま、麓で何匹か確認された程度ではなかろうか?
それは、素人が考えても“証明”とは云えない!
また日本のプロも、外国人の“論文”を頻繁に引用する。
アマチュアは苦労して外国人の“論文”を入手する気もないし、読む気もない。
だから、権威付けには手頃な作業なのだ。
それにしても、つまらない権威付けなどに浮き身をやつしていないで、
「自分で汗をながせ! と云いたい。
こんなインチキが罷り通るから、
蝶屋は正統派自然科学(数学、物理、化学、医学も)から馬鹿にされるのである。
日高敏隆氏は著書で、そのことを嘆いていたが自業自得という面もある。
そもそも論文に対する学会内での批判・反論・再反論がないのであるから!
多分、批判すれば自分も仕返しされることを恐れているのだろう。
それでは昇進に差し支える・・
蝶学は自然科学ではないなぁ!
④ 数理モデル、コンピュータ・シミュレーション
プロの著作に、数理モデルだのコンピュータ・シミュレーションなどという用語を見かける。
“数理モデルで、ある蝶の生態を解析した云々”
例えば、http://ecol.zool.kyoto-u.ac.jp/homepage/Ide/idejpn.htm
データも無しに生き物を論ずることが出来るのは創造主の御業である。
人間が“数理モデル”でやれることは、
・データを集める。
・データに基づいて自分の立てた仮説を確かめる。
(仮説を立てるのにコンピュータや数学が役立つこともあるだろう。)
・データを更に集めて、仮説を確かめる、或いは補強する。
ではなかろうか。
この作業で、データの整理にはコンピュータは有用ではあるが、
データ収集と洞察は依然として人間の仕事である。
データをコンピュータで生成してはなるまい。
これは、世を騒がす“データ捏造”である。
そもそも、変化自在の生き物の世界で、
数式による法則化なんぞ出来るものか!
ましてや、蝶屋の幼稚な数学で!!
⑤ 外国人の論文批判
十数年前、Hanski,Iという昆虫学者の論文を絶賛しているウェブサイトがあった。
私も入手して読んでみた。呆れかえった。以下。
P eunomiaという蝶(多分ヒョウモンモドキの仲間?)の移動を論じている。
約1km内の12パッチ(分断された蝶の生息エリア)間の移動を回帰分析で計算している。
・各パッチの個体数を面積の関数としている。
(個体数を食草の量の関数とし、更に食草の量を面積で置き換えたのだろう。)
・パッチ間移動量を、パッチ間距離と個体数とで説明している。
面積にしろ、パッチ間距離にしろ、時々刻々と変化しているだろう。
どうやって計算するのだろう?
特にパッチ間距離の場合、中間の草の種類、繁茂の程度を反映させるとすると、
殆ど計算のしようがないと思うが。
・なによりも呆れかえったのは、
「パッチ数は9以上をrecommend(推奨)する」だって!!
どういう意味かわかるだろうか?
計算すべきパラメータ数とパッチ数との関係を述べているのである。
パラメータ数に比して、パッチ数が或る程度大きくなければ、
オーバーフィッティングや0割が起きて無意味であると云っているのである。
生物学的見地からの要請ではない。単に、計算上の都合を云っているのである。
これは馬鹿げた要請である。
最初は、蝶は一つのエリアを飛び回っている。
やがて、人間の介入によってエリアは分断されパッチが増えていく。
二つ、三つ、・・・ 五つ・・九つ・・ やがて蝶は姿を消す
これを無念の思いで調べるのが、研究者の仕事ではないか。
こんな愚にもつかない要請を、最初から堂々とrecommendするな!
それじゃ、パッチ数、三つ、四つの時はどうするのだ!!
どう見ても、こっちのケースの方が生態学的にも重要ではないか!!
全く、別個のnewモデルを創出する御積りか?
日高敏隆氏は“外人は論文としての形式さえ整っていれば強引に出してくる。”
と零していたが、日本人でも同じではなかろうか。
日本人の場合は、更に外国人のエエ加減な論文をアリバイ作りに利用するから
一層たちが悪いような気がする。
⑥ 統計学
私も、蝶をカウントするけれどこれは統計データではない。
単に、データの要約・整理である。
年次間比較、月別比較、地域間比較に都合が良いからであって、
難しい統計処理などしていない。出来ないからだ。
蝶の学習能力に関して、「有意性云々」なる統計学用語を乱発している論文があった。
果たして蝶学に「有意性」なる「統計学用語」は適用出来るのか?
思いつくままに駄弁を弄する。
人口5万人の都市の性別を、サンプル数、1千人で調査する。
男性調査員は面倒くさがって手っ取り早く、手近の男子校からサンプリングするとする。
男=100%となる。
女性調査員は手っ取り早く女子校からサンプリングするとする。
女=100%となる
これがインチキであるとすぐ分かるのは、我々は人間の社会を良く知っているからである。
しからば結果が、男=80%、女=20%ならどうか?
限りなくインチキ臭いけれど、事実かも知れない。
実際、そういう集団もあるわけだから。
かくの如く、統計学というのはそれだけでは難しいというかインチキと判別出来ないというか。
だから、それ以外の知識も動員して判断するわけだ。
然らば、蝶の場合はどうか?
我々は蝶の集団について何も知らないに等しいのだから、
或る命題について、「有意であった、なかった」とか主張されても
判断のしようがない。
例えば、「蝶に学習能力ありの仮説」は有意であったと主張されたとする。
多分、そのサンプル集合は統計学的には全く無意味であろう。
かと言って、それ故にその仮説は成り立たないとも云えない。
結局、
「誠実に調査をやったのかどうか」
に帰するのである。
その誠実度(汗、人柄、博識etc)から主張の真偽を判断するしかない。
(そうすると、あまり自然科学らしくなくなってくるが・・)
いろいろな論文に、例えば10個くらいの点と直線の絵を見かける。
データの散布図と回帰直線の積りらしい。
こんなものは、街頭の落書きに如かず なのだ。
但し、自分の手抜き観察、インチキ観察のための弁明、権威付けには使える。
こういう実話があるそうだ。
ある公害裁判で、会社側の統計学者が
「排水は、公害病と因果関係があるとは云えない」
原告側が怒って、
「それじゃ、排水毎日送るから飲め!!」
統計学者は消耗したそうだ。
蝶学にはそのような厳しさは全くないから、
気楽にダボラを吹けるのである。
〔戻る〕
占有域以外では、こういう光景も良く見かける。
路上は好みの場所である。
但し遺伝子に組み込まれた占有域ではない。
だから派手な喧嘩はしない?
⑧ 総括
かくの如く、集結し始めてからやがて姿を消す迄、いずれかの個体が占有域に陣取っている。
占有個体は、そのポイントを死守しようとするが、頻繁に入れ替わっているようでもある。
その間、占有域で交尾行動を目撃したことはない。
どうみても、占有行動は雌確保の為ではない。
“その場所”が気に入っているからと考えた方が良さそうだ。
“その場所”は極狭い範囲である。
ちょっと離れると見向きもしない。
成蝶の行動は、採餌、交尾、産卵、休息、睡眠から成るが、
占有行動は、気に入った場所での休息と思う。
そこを確保するためには、闘いも辞さず というのが可笑しい。
5 ヒメアカタテハの発生回数(別サイトへ移行)
6 終わりに
蝶界への疑問を脈絡もなく考えている。
例えば、
・蝶道 ・占有行動 ・ヒルトッピング ・交尾飛翔形式
・野外での観察と実験室での“研究” ・標本の所有者と“研究者”
・1化と遺伝的1化とダラダラ1化
など。
今回は、ヒメアカタテハの占有行動と交尾との関係について述べた。
他に考えていることを述べてみる。
① アオバセセリの占有行動
アオバセセリについて述べた論文を読んだ。
それによれば、
観察期間中、縄張りでの交尾を1回目撃した。
他にもいろいろの状況証拠を揃えた。
よって、
占有行動―>交尾
と結論づけた
ということらしいが、私にはさっぱり分からない。
・0回目撃なら氏は自説を撤回するのだろうか?
然らば、0回と1回には天と地程の違いがあるが
そんな違いはあるのだろうか?
・例えば、
「アオバセセリとスミナガシは同じ位の大きさだから
雌と誤認して執拗に追いかける」
なる観察がある。
どうして、これらが状況証拠になるのだろうか?
アオバセセリの占有行動も交尾とは関係がないだろう。
「山頂に惹かれて集まってくる」のだと思う。
② 蜂の話題
ミツバチの生態について書かれている著書、ウェブサイトは多い。
配偶行動について取りあえず私が紹介する。(不正確かも知れない)
*************************************
・春、雄バチ群が羽化する。1日数回、巣を出入りする。
交尾場所を探索、確認しているらしい。
・そのうち、女王バチ候補が羽化する。
雄バチは、同じ巣の女王バチ候補には全く関心を示さない。
つまり、近親交配を忌避しているのである。
・各巣から出陣した雄バチは、特定の交尾場所に集結している。
女王バチ候補が性フェロモンを振りまきながら現れると、
目敏く見つけた雄群が黒い塊となって女王バチ候補を追いかけ
運の良い雄が交尾をする。
・雄バチは1回の交尾で死ぬが、女王バチ候補は、7~8回交尾をするらしい。
(ということは、子どもの働きバチの父親は7~8匹いることになる)
・問題は交尾場所である。白昼、黒い塊が唸りを上げて飛び回っている。
高さ:10~40m 直径:50~200m
極普通に飛んでいるミツバチなのだから、
素人でも交尾場所は発見出来そうだが、これが相当の難問らしい。
各国の発見例が紹介されていて、日本では未発見となっていた。(或る著書では)
その後、日本でもハイテク機器を駆使して発見されたようだ。
・もう一つの難問は、《その交尾場所が毎年同じ場所であること》である。
ハチが集合場所を探す手掛かりは今のところ、
目立つ目印や特徴的な地形、つまり視覚的なもの とされている。
・トウヨウミツバチ(亜種ニホンミツバチ)、スズメバチについても
いろいろな研究が報告さている。
***************************************
読んでいて、私のような素人でも興奮するのである。
目下、二つの点が気になっている。
一つ目は、ハチは今や家畜であるということである。
養蜂家としては、
「働きの良い巣をつくりたい。
女王バチが勝手に外に出かけていって、
体力はあるが怠け者のオスと勝手に交尾してもらっては困る。」
だから、近親交配は避けるというハチが守ってきたルールは
養蜂家によって破られているようなのである。
それがハチの反乱、即ち
「もう、かんべんしてくれ!」ということで、
働きバチが巣に帰ってこない--->巣の崩壊
をもたらしているのではないだろうか?
(実際はどうなっているか分からないけれど)
二つ目は、以下の指摘である。
「ハチが集合場所を探す手掛かりは今のところ、
目立つ目印や特徴的な地形、つまり視覚的なものである。」
つまり、「このような光景へ惹かれること」が遺伝子に組み込まれていると云うことだ。
毎年産まれてきた雄や女王バチ候補が同じ場所に集まってくるのだから、
こう考えざるを得ないということだろう。
こういう論法は、蝶の行動(一部の種の一部の個体の行動)にも当てはまるのではなかろうか。
例えば、山登り(ヒルトッピング)、占有行動。
(但し蝶の場合は、このことと配偶行動とは関係がないと私が思う所以は後段で。)
③ 蝶の話題
然らば、蝶の場合はどうであろうか?
ヒルトッピング、レック型、非レック型なる“専門用語”がある。
これらに関する記述は、いろいろな著書やウェブサイトで見られるが、
例えば、https://www.pteron-world.com/topics/ecology/mating.html
アメリカの学者シールズは、
「山に集まって来る雌は交尾していない個体で、山頂で交尾して山を下る。」ことを“証明した”らしい。
(因みに、その後多用されている「レック」というのは
蝶が交尾のために集まってくる場所をさすらしい。シールズが命名したのかな?)
アメリカ人がアメリカの山でどういう調査をしたのか知らないけれど、
そのような学説があるのなら、日本人は日本で確かめてみれば良いではないか。
私は子どもの頃から、いろいろな山に登ったけれど、
交尾が活発に行われている光景などまるで思い出せない。
多数の蝶が集まって来る極せまい山頂にも何度も登ったけれど、
そもそも交尾そのものが全く思い出せないのである。
(殆ど雄だけだったような気がする。)
往時は交尾行動に特別に関心があったわけではないが、
この十数年は、岩手県の山、東京周辺の山を調べてみた。
シールズのように「入り鉄砲に出女を調べる関所の役人」のような熱心さで
調べたわけではないが、それでもそのような事例は全く目撃出来なかった。
ウェブサイトを見ても、
「集合場所では雌は目撃できなかった、交尾は目撃出来なかった」
という報告ばかりである。
少なくとも、《蝶の「占有場所」はミツバチの「交尾空間」と同じ》なる報告は全くないのである。
私は、シールズの“証明”は、アメリカにあっても“false”であると思っている。
頂上でマーキングした雌がたまたま、麓で何匹か確認された程度ではなかろうか?
それは、素人が考えても“証明”とは云えない!
また日本のプロも、外国人の“論文”を頻繁に引用する。
アマチュアは苦労して外国人の“論文”を入手する気もないし、読む気もない。
だから、権威付けには手頃な作業なのだ。
それにしても、つまらない権威付けなどに浮き身をやつしていないで、
「自分で汗をながせ! と云いたい。
こんなインチキが罷り通るから、
蝶屋は正統派自然科学(数学、物理、化学、医学も)から馬鹿にされるのである。
日高敏隆氏は著書で、そのことを嘆いていたが自業自得という面もある。
そもそも論文に対する学会内での批判・反論・再反論がないのであるから!
多分、批判すれば自分も仕返しされることを恐れているのだろう。
それでは昇進に差し支える・・
蝶学は自然科学ではないなぁ!
④ 数理モデル、コンピュータ・シミュレーション
プロの著作に、数理モデルだのコンピュータ・シミュレーションなどという用語を見かける。
“数理モデルで、ある蝶の生態を解析した云々”
例えば、http://ecol.zool.kyoto-u.ac.jp/homepage/Ide/idejpn.htm
データも無しに生き物を論ずることが出来るのは創造主の御業である。
人間が“数理モデル”でやれることは、
・データを集める。
・データに基づいて自分の立てた仮説を確かめる。
(仮説を立てるのにコンピュータや数学が役立つこともあるだろう。)
・データを更に集めて、仮説を確かめる、或いは補強する。
ではなかろうか。
この作業で、データの整理にはコンピュータは有用ではあるが、
データ収集と洞察は依然として人間の仕事である。
データをコンピュータで生成してはなるまい。
これは、世を騒がす“データ捏造”である。
そもそも、変化自在の生き物の世界で、
数式による法則化なんぞ出来るものか!
ましてや、蝶屋の幼稚な数学で!!
⑤ 外国人の論文批判
十数年前、Hanski,Iという昆虫学者の論文を絶賛しているウェブサイトがあった。
私も入手して読んでみた。呆れかえった。以下。
P eunomiaという蝶(多分ヒョウモンモドキの仲間?)の移動を論じている。
約1km内の12パッチ(分断された蝶の生息エリア)間の移動を回帰分析で計算している。
・各パッチの個体数を面積の関数としている。
(個体数を食草の量の関数とし、更に食草の量を面積で置き換えたのだろう。)
・パッチ間移動量を、パッチ間距離と個体数とで説明している。
面積にしろ、パッチ間距離にしろ、時々刻々と変化しているだろう。
どうやって計算するのだろう?
特にパッチ間距離の場合、中間の草の種類、繁茂の程度を反映させるとすると、
殆ど計算のしようがないと思うが。
・なによりも呆れかえったのは、
「パッチ数は9以上をrecommend(推奨)する」だって!!
どういう意味かわかるだろうか?
計算すべきパラメータ数とパッチ数との関係を述べているのである。
パラメータ数に比して、パッチ数が或る程度大きくなければ、
オーバーフィッティングや0割が起きて無意味であると云っているのである。
生物学的見地からの要請ではない。単に、計算上の都合を云っているのである。
これは馬鹿げた要請である。
最初は、蝶は一つのエリアを飛び回っている。
やがて、人間の介入によってエリアは分断されパッチが増えていく。
二つ、三つ、・・・ 五つ・・九つ・・ やがて蝶は姿を消す
これを無念の思いで調べるのが、研究者の仕事ではないか。
こんな愚にもつかない要請を、最初から堂々とrecommendするな!
それじゃ、パッチ数、三つ、四つの時はどうするのだ!!
どう見ても、こっちのケースの方が生態学的にも重要ではないか!!
全く、別個のnewモデルを創出する御積りか?
日高敏隆氏は“外人は論文としての形式さえ整っていれば強引に出してくる。”
と零していたが、日本人でも同じではなかろうか。
日本人の場合は、更に外国人のエエ加減な論文をアリバイ作りに利用するから
一層たちが悪いような気がする。
⑥ 統計学
私も、蝶をカウントするけれどこれは統計データではない。
単に、データの要約・整理である。
年次間比較、月別比較、地域間比較に都合が良いからであって、
難しい統計処理などしていない。出来ないからだ。
蝶の学習能力に関して、「有意性云々」なる統計学用語を乱発している論文があった。
果たして蝶学に「有意性」なる「統計学用語」は適用出来るのか?
思いつくままに駄弁を弄する。
人口5万人の都市の性別を、サンプル数、1千人で調査する。
男性調査員は面倒くさがって手っ取り早く、手近の男子校からサンプリングするとする。
男=100%となる。
女性調査員は手っ取り早く女子校からサンプリングするとする。
女=100%となる
これがインチキであるとすぐ分かるのは、我々は人間の社会を良く知っているからである。
しからば結果が、男=80%、女=20%ならどうか?
限りなくインチキ臭いけれど、事実かも知れない。
実際、そういう集団もあるわけだから。
かくの如く、統計学というのはそれだけでは難しいというかインチキと判別出来ないというか。
だから、それ以外の知識も動員して判断するわけだ。
然らば、蝶の場合はどうか?
我々は蝶の集団について何も知らないに等しいのだから、
或る命題について、「有意であった、なかった」とか主張されても
判断のしようがない。
例えば、「蝶に学習能力ありの仮説」は有意であったと主張されたとする。
多分、そのサンプル集合は統計学的には全く無意味であろう。
かと言って、それ故にその仮説は成り立たないとも云えない。
結局、
「誠実に調査をやったのかどうか」
に帰するのである。
その誠実度(汗、人柄、博識etc)から主張の真偽を判断するしかない。
(そうすると、あまり自然科学らしくなくなってくるが・・)
いろいろな論文に、例えば10個くらいの点と直線の絵を見かける。
データの散布図と回帰直線の積りらしい。
こんなものは、街頭の落書きに如かず なのだ。
但し、自分の手抜き観察、インチキ観察のための弁明、権威付けには使える。
こういう実話があるそうだ。
ある公害裁判で、会社側の統計学者が
「排水は、公害病と因果関係があるとは云えない」
原告側が怒って、
「それじゃ、排水毎日送るから飲め!!」
統計学者は消耗したそうだ。
蝶学にはそのような厳しさは全くないから、
気楽にダボラを吹けるのである。
〔戻る〕
ティータイム(その1) 「不思議なシマヘビの物語 (野川で出会った“お島”)」
ティータイム(その2) 「ミノムシ 《皇居外苑北の丸公園の蓑虫》」
ティータイム(その3) 「ゴイシシジミ賛歌」
ティータイム(その4) 「空飛ぶルビー、紅小灰蝶(ベニシジミ)」
ティータイム(その5) 「ヒメウラナミジャノメの半生(写真集)」
ティータイム(その6) 「蝶の占有行動と関連話題」
ティータイム(その7) 「ヒメアカタテハの占有行動」
ティータイム(その8) 「オオウラギンヒョウモン考」
ティータイム(その9) 「ヒメウラナミジャノメの謎」
ティータイム(その10) 「コムラサキ賛歌」
ティータイム(その11) 「散歩しながら動物行動学を学ぶ」
ティータイム(その12) 「”お島”ふたたび」
ティータイム(その13) 「オオウラギンヒョウモン考(再び)」
ティータイム(その14) 「謎の蝶 ヒメアカタテハ」
ティータイム(その15) 「我が隣人 ヒメアカタテハ」
ティータイム(その16) 「オオウラギンヒョウモン考(三たび)」
ティータイム(その17) 「姿を顕さない凡種、クロヒカゲ」
ティータイム(その18) 「微かに姿を顕したクロヒカゲ」
ティータイム(その19) 「不可解な普通種 ヒメジャノメ」
ティータイム(その20) 「散歩しながら動物行動学を学ぶ ― 蝶の知的生活―」
ティータイム(その21) 「オオルリシジミを勉強する」
ティータイム(その22) 「集結時期のヒメアカタテハを総括する」
ティータイム(その23) 「毒蛇列伝」
ティータイム(その24) 「東京ヘビ紀行(付記 お島追想)」
ティータイム(その26) 「ヒメアカタテハの越冬と発生回数」
ティータイム(その27) 「鳩山邦夫さんの『環境党宣言』を読む」
ティータイム(その28) 「蝶の山登り」
ティータイム(その29) 「蝶の交尾を考える」
ティータイム(その30) 「今年(2019年)のヒメアカタテハ」
ティータイム(その31) 「今年(2019年)のクロヒカゲ」
ティータイム(その32) 「蝶、稀種と凡種と台風と」
ティータイム(その33) 「ルリタテハとクロヒカゲ」
ティータイム(その34) 「ヒメアカタテハ、台風で分かったこと」
ティータイム(その35) 「「蝶道」を勉強する」
ティータイム(その36) 「「蝶道」を勉強する 続き」
ティータイム(その37) 「ミツバチを勉強する」
ティータイム(その38) 「「蝶道」を勉強する 続き其の2」
ティータイム(その39) 「里山の蝶」
ティータイム(その40) 「岩手の蝶 ≒ 里山の蝶か?」
ティータイム(その41) 「遺伝子解析、進化生物学etc」
ティータイム(その42) 「今年(2020年)の報告」
ティータイム(その43) 「徒然なるままに 人物論(寺田寅彦、ロザリンド・フランクリン、木村資生、太田朋子)」
佐藤特許事務所(世田谷区太子堂)のサイト 〔トップページへ戻る〕
|