
![]()
![]()

�i1���N�O�̉H�т̉��B���̂��̂������̂��̂��͕s���B���E�Ώ̂Ȃ̂Ŕ��ėp�ł͂Ȃ��悤���B�j
���{�����I�s��6��u�����̐i���Ɛ�Łv�ł��q�ׂ����A���͋������`������������ׂĔ����Ă���B�t�ɒ����`��������i�P�����A�H�сA�����j����������������������Ă���B�����ƒ��̋�ʂ͓���A�Ƃ�����������s�\�ł���B�܂�A���͋����̈ꕔ�A���������ƍl���邱�Ƃ��ł���̂ł���B�܂�A�����͐�ł��ĂȂ���X�̂������ł��������Ă���̂��B
�ł͋����͂ǂ̂悤�ȓ������ǂ��Ē��ɂȂ����̂��낤���B���̋^��ɂ��Ă��V�ق̓W���̓q���g��^���Ă����B
�P�D���ւ̓�
�y�h���}�G�I�T�E���X�ށF�o���r���v�g���H�z
 �@�@
�@�@
�i���ʂ���B�e�j�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�i���ʂ���B�e�j
�y�n�c���F�A���P�I�v�e���N�X�z
 �@�@
�@�@
�i�RD�̑g�ݗ��ĕW�{�j�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�i�H�т̍��Ղ��L���ȃx�������W�{�A���قɓW���j
�y���̍��i�z�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�y�����ƒ��z
 �@�@
�@�@
�i�����ł����Ԃ̃n�g�̍��i�j�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�i��O�������ʼn������A�i���̗����A�����Č����悤�ɍH�v����Ă���j
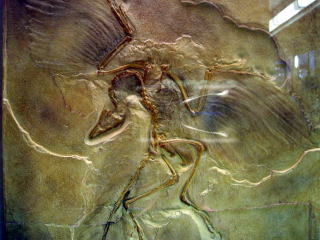
�V�قɂ͋������n�c�����n�g�ƘA�����č��i���r�ł���悤�ɓW������Ă���B�ʐ^�����Ă����������������ɂȂ�Ǝv�����A�����i�h���}�G�I�T�E���X�j�Ǝn�c�̒����i�͋����قǎ��Ă���B�܂��n�c���ƃn�g�̍�������ׂĂ݂��
�ȉ��̂悤�ȓ��������邱�Ƃ��ł���B
�E���̍b�̍����܂Ƃ܂��Ă���B�i���̓������h���}�G�I�T�E���X�����e�B���m�T�E���X�ɂ����ʁj
�E���̍��̏�ɍ����i�H�����ۂɃo�l�ɂȂ鍜�j������B�i������h���}�G�I�T�E���X�ނ�e�B���m�T�E���X�Ƃ����ʁj
�E���̍����ۂ��A�r������܂肽���߂�B�i������h���}�G�I�T�E���X�ނƂ����ʁj
�E���̊߂����������r���������ɍL���邱�Ƃ��ł���i������h���}�G�I�T�E���X�ނƂ����ʁj
���̂��A���ꂪ�d�v�����`�͈Ⴄ����{�I�Ƀn�g�ɂ��鍜�i���̍��A���̍��Ȃǁj�͂������������A�`�͑傫���Ⴄ����ʂ�n�c���������Ă���B���̂��Ƃ͋t�ɒ����K���⎕�Ƃ����������Ǝn�c���̋��ʕ������n�g�ł͎����Ă�����̂́A�n�c���ƃn�g�͌`�͈Ⴄ�����ʂ̃p�[�c�Ō`������Ă���̂ł���B�������A���̃p�[�c�͋����������Ă���̂ł���B�܂��n�g���������`������������ׂĎ��Ă��邱�Ƃ͐�قǏq�ׂ��Ƃ���ł���B
�܂蒹�͋���ԂƂ����K���̂��߂ɋ����̎��p�[�c�̌`��ό`�����A�K�v�̂Ȃ����̂͂��Ă��̂ł���B���̂悤�Ȃ��Ƃ���A���͔�Ԃ��߂ɐi�����������Ƃ�����̂ł���B
�Q�D�߂͌�闃�̗��j
���̗��͋����O�����i���������̂ł���B
��������Ε�����Ǝv�����A���͔�ԂƂ�����̂̉��ɑ傫���L���A�����������㉺�ɓ������ĉH�����A�n��ł͗���܂肽���ށB������O�̂��Ƃ������̂悤�ȓ�������邽�߂ɂ͌��̊߂Ǝ��̊߂ɂ��Ȃ�̎��R�x���K�v�ɂȂ��Ă���B
������ɉH�������߂ɂ͊߂̐i�����K�v�ł������B�����Ă��̐i���͋�����������p�������̂ł������̂��B
�y���̊߁z
 �E�̎ʐ^�͋����̎��̊߂̔�r��\���Ă���B
�E�̎ʐ^�͋����̎��̊߂̔�r��\���Ă���B
��O���ŏ��̋����w�������T�E���X�̎��B�����f�B�m�j�N�X�̎��ɂȂ�B�f�B�m�j�N�X���̂̏Љ�͂��Ƃŏڂ����s�����A�f�B�m�j�N�X��������h���}�G�I�T�E���X�ނ��g���G�h���Ƃ������b�r�ނ̓}�j���v�g���ނƕ��ނ���A���ނɌn���I�Ɉ�ԋ߂������Ƃ���Ă���B
���̓W���ł͗��҂̎��̓�����������Ă���B
�悸�w�������T�E���X�̓��������A�ނ�̎��͏㉺�����ɂ��������Ȃ��B�܂菵���L�̂悤�ɂ�����������Ȃ��̂��B�ނ�̎��͒P���ɂ��̂��ォ�牟������Ƃ��������������ł��Ȃ������̂ł���B
����A�f�B�m�j�N�X�ł͎��̍����i�ʐ^�ʼn��F���Ȃ��Ă��镔���j�ۂ��Ȃ��Ă���A���̂������ŏ㉺���E�A�܂蔼����ɂɎ��R�ɓ��������Ƃ��ł���B����ɂ���Ĕނ�͒���������ނ悤�ɘr���Ȃ��邱�Ƃ��ł����B�����炭����͊l���������I�ɂ���A�������肷��̂ɗL���������̂��낤�B�ӊO�ƁA�������Ƃ��ċ@�\���邽�߂̗v���͋������ʂȖړI�Ŕ��B�����Ă����̂�������Ȃ��B
�y���̊߁z


�i�E�F�f�B�m�j�N�X�̌��߁A���F�e�B���m�T�E���X�̌��߁w�B�e�ꏊ�F�_�C�m�\�A�t�@�N�g���[�x�j
��̎ʐ^�̓f�B�m�j�N�X�ƃe�B���m�T�E���X�̌��߂̔�r�ł���B
�f�B�m�j�N�X�͌��̊߂̖ʂ��傫���A���̓I�ɂȂ��Ă��āA���Ɍ����Ă��Ă���悤�Ɍ�����B���Ȃ�r���㉺���E�ɔ�����Ȏ��R�x�̍����������ł��邱�Ƃ��\�z�ł���B����͗����H�������邽�߂ɂ͕K�v�ȓ������B
����A�e�B���m�T�E���X�̌��߂͎ߌ��ɂ��Ă���B�ߖʂ��������B�f�B�m�j�N�X�Ɣ�r����Ɨ\�z����铮���͈̔͂��������Ǝv����B�����炭���ɂ͂�����x�������A�O�≡�̓����͐������ꂽ���낤�B
�R�D���͂ǂ����Ĕ�̂��H
�����܂łŒ��̉H�����ɕK�v�Ȋ߂̓����́A���ނɋ߂������ł��łɎ�������Ă������Ƃ��������Ă����������Ǝv���B�H�т��܂��ŋ߂̉H�ы����̔��@�ŁA��Ԃ��ƂƂ͊W�Ȃ������̓����Ƃ��Ĕ��B���Ă������Ƃ��������Ă����B������Ԃ��Ƃ͉H�����ƉH�тƂ����ǂ��炩�ƌ����ƕʁX�ɐi�����Ă����v�f�����킳�����Ƃ��ɕ����I�ɐ������ƌ�����̂ł���B�܂�A���͍ŏ������Ԃ��Ƃ�ڎw���Đi�������̂ł͂Ȃ��A���܂��ܔ�Ԃ��Ƃ��ł��A���ꂪ�����c��̂ɗL���������Ԕ\�͂B�������ƌ������Ƃł���B
�y�����P���������ƉH�т̋N���z
�H�тɂ��ẮA���Ƃ��Ə��^�̃R���G�T�E���X�ނŔ��B�������Ƃ��������Ă���B
�����͍P���������������ǂ����ɂ��ẮA�c�_��������Ă���B������������A�a���������������͍P�������Ƃ��Ă͕s���S�Ȑ������������̂�������Ȃ��B�O��I�̌��������̒��ŃG�l���M�[���ʂɏ����P�����̊l���͗L���Ȑi���Ƃ͂����Ȃ����낤�B�����Ŋ�����������A�}���Ő������邽�߂Ɏq������ɍ�����ӂ��ێ����邽�߂ɍP�����������Ƃ͗L����������������Ȃ��A��������������������G�l���M�[������邾���̍P�����͎ז��ł͂Ȃ������̂��낤���H������A�ŏ����̋��������͊��S�ȍP�������łȂ����������߂ɔɉh�̃`�����X�����ɈႢ�Ȃ��B�q���̎���͐����̂��ߍ�����ӗ��������Ă������̂́A��l�ɂȂ�Ƒ�ӂ�������ω������Ƃ��Ȃ����x���A�������͋��剻�ɂ���Ċ����P�������ɂȂ����̂ł͂Ȃ����낤���B
�������A�����̈ꕔ�ɂ͎q������̍P���������̂܂܈ێ������푰�������̂ł͂Ȃ����낤���B���̑�\�I��̓R���G�T�E���X�ނł͂Ȃ����낤���B�ނ�͉H�тő̂�ۉ����邱�ƂŖ��ʂȃG�l���M�[�����}���悤�Ƃ����ƍl������B
�y�Q�l�z
�i�����P�����j
�ω������ł��̂�����Ȃ��̂͑̉��̕ω����قƂ�ǂȂ��B�����݂̂����ƕ��C�̓��ł͗ʂ̑������C�̂ق������߂ɂ����̂Ɠ��������ł���B���������ł̕�����₷����Ƃ��Ă��I�T�K���i�S���Q���̊C�T�j�͋��̂䂦�ɑ��̊C�T�ɂ���ב̉��ω������Ȃ��A�x�[�����O�C��k�C�A�o�����c�C�Ȃǖk�Ɍ��ɋ߂��C��ł������ł���B�I�T�K����������ȋ��������̌̂ɑ̉�����肾�����ƍl������B
�i�ω������ƍP�������F���嗳�r�ނ̏ꍇ�j
���Ȃ̋��ȏ��ł͍P���������ω����������D��Ă���悤�ɏ����Ă��邪�A�K�����������ł͂Ȃ��B
���܂��܌��݂̒n���ł͍P�������̂ق��������₷�����Ȃ��߁A�ɉh���Ă��邾���ł���B�ω��������P���x�������D�ꂽ�ʂ����Ȃ肠��̂��B�Ⴆ�A�ω������ɂ͍P�������ɔ�אH���̗ʂ����Ȃ��Ă��ނƂ��������b�g������B����͍P���������̉����ێ����邽�߂ɃG�l���M�[��K�v�Ƃ��邽�ߐH���̗ʂ𑝂₷�K�v������̂��B����ɂ͕ω������͑̏d�������ł���P�������̂P/�P�O�ł��ނƂ����B
����������A�Ƃ��ɑ�^�̗��r�ށi�J�~�i�����j�ɓ��Ă͂߂čl���Ă݂悤�B
�O��I�`�W�����I�`�����I�O���ƒ�����̑唼�̎���ɗD���ł������V�_�A���A���q�A���i�j�t���Ȃǁj�͔�q�A�������h�{�����Ⴂ�Ƃ����B�����̋��剻�̈�̃s�[�N�̓W�����I����ł���A���̎���͉h�{���̒Ⴂ�V�_�A���A���q�A�������Ȃ��ƌ����ł������B�����Ƃ���q�A�������̎��ォ��킸���ł͂��邪���݂����B�������A���嗳�r�ނ��猩��Ȃ������R�̂��̂������B
�����A�u���L�I�T�E���X���Z�C�X���T�E���X�̂悤�ȋ��嗳�r�ނ��P�������������ƍl������A�ނ�͋��̂��ێ����邽�߂ɕω������̏ꍇ��
�P0�{�Ƃ����c��ȐA����������ɈႢ�Ȃ��B�܂��h�{�����Ⴂ���߂��̗ʂ͔��[�ł͂Ȃ��������낤�B�����H�ׂ邽�߂����Ɏ��Ԃ��₷���ƂƂȂ����̂��낤���A�ق�Ƃ��ɊԂɍ������̂��낤���H�������A���L�^������Ɠ����n��̒��ɑ����̎�ނƁA���Ȃ�̌̐������݂����悤�ł���B�܂�ނ炪�P�������������琶�Ԍn�ɂ��Ȃ�̈��͂ɂȂ����̂ł͂Ȃ����낤���H�������r�ނ��P�������Ȃ�^��_�������B�������A�ނ�ɂƂ��čP�������L���ȓK���Ƃ͎v���Ȃ��B
�ނ���A�ނ�͊�{�I�ɏ��H�ł��ޕω������ŁA���̂ɂ���đ̉������ɕۂ����P�������ł������Ƃ�����ǂ����̐����낤�B���Ȃ��H���Ŋ����Ȋ������ł����̂ł����ƍl������B���Ԍn�Ɉ��͂������Ȃ����x�ɑ��l�Ȏ�ƌ̐����ێ��ł����̂ł͂Ȃ����낤���B
���̂悤�ɍl���Ă݂�ƁA�K�����P���������L���ƌ����킯�ł͂Ȃ��悤�ł���B
�y�]�k�F�n�����嗤�ʂ̋����P���������z
�]�k�����A���������͕s���S�ȍP�������������Ƃ��Ă��A���ǂ̂Ƃ����r�I�ɑ����i�K�ŋ��剻�������r�ނ������đ����̋������P�������������̂ł͂Ȃ����낤���B���^�̋�������i�����Ă�����̂̑����͑c��̒i�K�ōP�������l�����Ă��܂��������m��Ȃ��B�����I�̖k�����̎�ȋ����̌n���͏��^��������i�������B������A�����I�̖k���������̋����͍P��������������������Ȃ��B
�Ⴆ���g���P���g�v�X�Ȃǂ̊p���B�ނ�̑c�擮�����v�V�b�^�R�T�E���X�̂悤�ȏ��^�����������B�v�V�c�^�R�T�E���X����H�т̂悤�ȍ��Ղ����������ƌ�����������B�g���P���g�v�X�ɉH�т��������Ƃ͎v���Ȃ����A�ނ炪�����悭�ł���V�X�e���B�������͕̂K�v�Ƃ����G�l���M�[�邽�߂̓K���������̂����m��Ȃ��B�n�h���T�E���X���̃f���^���o�b�e���[���������R�ł̓K����������Ȃ��B
����A��r�I�����ɑc�悪��^�������씼���̋����F���r�����P���g�T�E���X���Ȃǂ͍P�������ł͂Ȃ��������̂����������̂�������Ȃ��B
���̂悤�ɋ������P���������������ǂ����͑c�悪��^��������������ɂ���ĈقȂ�\��������B�܂��A�����I�ɖk�����Ɠ씼���ŋ����̎�ނɑ傫�ȈႢ��������̂��A�n���I�Ȓf�₾���ł͂Ȃ��P�������̂ق����L�����ǂ����Ƃ������̈Ⴂ���傫�������̂ł͂Ȃ����B���̖T�Ƃ��āA�씼���ł���Ɍ��ł����G���i�T�E�����͂��߂Ƃ��鏬�^�̃q�v�V���z�h���ނ��ɉh�������Ƃ���������B�ނ�͍P���������������̂ɓ�Ɍ��Ŕɉh�ł����̂�������Ȃ��B
�y�H�����̋N���z
�b�����ł��܂������A�H�т̒a���͑̉����߂Ɩ��ڂȊW������̂��낤�B
�ł͉H�������ł���߂̍\���͉����琶�܂ꂽ�̂��낤�B����ɂ͒��ƃ}�j���v�g���ނ̑c��Ɋւ���2�̉������z�肳���B
�@�l����߂܂��邽�߂ɉ��͈͂��傫���߂����B�����B
�A����Ȃlj��͈͂��傫���߂��������ق����������ɏZ��ł���
�@�Ɋւ��Č����ΐ�قnj����Ƃ����e�B���m�T�E���X���A���T�E���X�Ȃǂ̑��̕ߐH�����ł͂���قNJ߂̉��͈͂�K�v�Ƃ��Ȃ��������Ƃ��l������ЂƂ����͂��ア�ƌ��킴��Ȃ��B����A�A�ɂ��Ă̓}�j���v�g���ނŎ��㐫���~�N�����v�g���̑��݂�����B�ނ�͌��r�ɂ��������������ƂŗL���ȋ����ł���B
�~�N�����v�g�����肪����Ɋߍ\���̐i�����l���Ă�A���ƃ}�j���v�g���ނ̑c��͎��㐫�Ŏ}��͂�A�����݂����߂ɉ��͈͂��傫���߂̕K�v�����������Ɛ����ł���B
�����炭�A�~�N�����v�g���͒��ƃ}�j���v�g���ނ̑c��̌`�����p���������������̂�������Ȃ��B
�܂�A���̐�c�ƂȂ鋰���̓~�N�����v�g���̂悤�Ȏ��㐫�̏��^�����ŁA��X�̐��E�̉��̂悤�Ɏ}��͂݁A���ɂ����݂����߂ɉ��͈͂̑傫�����⌨�̊߂��l�������B�܂��A�ނ�͏��^�ōP�������䂦�ɕۉ��̂��߉H�тB�����Ă����B�i������������f�B�X�v���C�ɂ��g�����̂�������Ȃ��j
�����Ĕނ�́A2�̌n���ɕ�����Ă������B����͕ۉ��p�̉H�тƉߓx�͈͂̑傫���߂𗘗p���H�����ɂ�����Ԃ��Ƃ��ł������㐶���ҁA�܂蒹�ނł���B��������͒n��ɉ���āA�h���}�G�I�T�E���X�ނ�g���G�h���Ƃ����������Ċl����߂炦��ߐH�����ɂȂ������́B���������^���v�g���̂悤�ȑ�^��ݏo���ȂǁA�����Ē��ނ�����^�����铹��I�B�}�j���v�g���ނ͂���Ӗ��_�`���E��y���M���̂悤�Ȓn��ɍ~�肽���̂悤�Ȃ��̂���������������Ȃ��B
�S�D���ޒa���̓�
�������܂��ߒ��̉���������܂ł݂Ă����B��x�A�������Ă݂悤�B
�@�@�@�@�O��I�s���S�ȍP�������i�q������͍P�����A������͕ω��A�����P�����j�Ƃ��ċ������X�^�[�g�����B
�@�@�@�A���^�̃R�G���T�E���X�ނ̌n���͎q������̍P�������l�ɂȂ��Ă��ێ����邱�ƂōP�������ɐi�������B
�@�@�@�B�R�G���T�E���X�ނ̌n���͕ۉ��̂��߉H�т�i���������B
�@�@�@�C�R�G���T�E���X�ނ̂������ƃ}�j���v�g���ނ̑c��ɂ����鋰���͎��㐶���𑗂��Ă���
�@�@�@�D�ނ�͎��㐶���ɓK�����邽�߁A�傫�����R�ɓ������E���̊߂��l�������B
�@�@�@�E�ނ�̌n���͈ȉ���2�n���ɕ����ꂽ�B
�@�@�@�@�@�E�n��ɍ~�葖�邱�ƂɓK�������ߐH�����ɐi�������}�j���v�g����
�@�@�@�@�@�E�H�тƊ߂̉��͈͂𗘗p������Ԃ��Ƃ��ł�����㐩��������
�������A��L�̉����ɂ͂������Ȗ�肪����̂��A�ȉ���������Ă݂悤�B
�y�n�����̖����z
�܂��A�ŏ��̏b�r�ނփ������T�E���X���N�_�ɒ��Ɏ���i���̓������Ă��炨���B
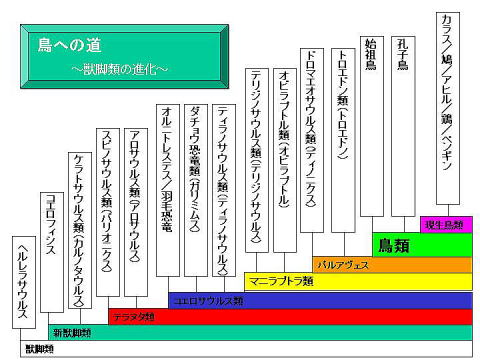
���Ȃ݂ɋ�̓I�Ȋe�X�e�b�v�ł̐i���̓����͂�������Q�Ƃ��Ă��������B
![]()
�オ����w�̎�@��p�����̓������r���ē���ꂽ�b�r�ނ̌n�����ł���B���n�I�ȏb�r�ނ��璹�܂łǂ̂悤�ȃX�e�b�v��Ői�����Ȃ���Ă����āA�����Ԃ̌n���̋߂���m��̂ɂ͔��ɗ��ɂ��Ȃ����}�ł���B
�������A���̐}�ɂ͑傫�Ȗ�肪���邱�Ƃ����C�Â����낤���H
�i�@�j�͂��̃O���[�v�̑�\�I�����ł���B�Ⴆ�g���G�h���͔����I����A�f�C�m�j�N�X�͔����I�O���A�I�r���v�g����
�����I����A�ŋߒ����ő��X�Ɣ��������H�ы��������͔����I�O���̋��������ł���B
����ŁA�n�c���̓W�����I����̃��[���b�p�ɐ������Ă����B
�����ŁA��̐}�̋�����\�Ő������Ă݂悤�B
�i������₷�����邽�ߕ\�̂��̈ȊO�Ŋe�n�����\���鋰�����ꕔ�lj�����B�j
| �O��I | �W�����I | �@�����I | ||
| ��� | �O�� | ��� | �O�� | ��� |
| �w�����T�E���X �R�G���t�B�V�X |
�f�B���t�H�T�E���X �i�P���g�T�E���X�ށj |
�P���g�T�E���X �i�P���g�T�E���X�ށj �A���T�E���X �I���j�g���X�e�X �n�c�� |
�J���m�^�E���X �o���I�j�N�X �H�ы��� �f�B�m�j�N�X �E�q�� |
�K���~���X �e�B���m�T�E���X �e���W�m�T�E���X �I�r���v�g�� �g���G�h�� |
��̕\������Ɗe�����������Ă������オ�n�����Ƃ͖��W�ł��邱�Ƃ������ł���Ǝv���B
�܂�S�[���ɐݒ肵���n�c��������̎���A�܂蔒���I�ɂȂ��Ē��ɋ߂������������o������̂ł���B
���̖������ǂ���������悢�̂��낤�H
����A�P���g�T�E���X�ނ̂悤�Ƀf�B���t�H�T�E���X���P���g�T�E���X���J���m�^�E���X�Ǝ���������ăO���[�v���̑�\�I����������������Ă���������B
�y�X�Ȃ鍬���F�v���g�A���B�X�z
��̐}�̖�������ɂ�₱�������鉻���k�Ă̎O��I����i2��5000���N�O�j�̒n�w�Ō������Ă���B���ꂪ�v���g�A�r�X���B2��5000���N�O�ƂȂ�ƁA�R�G���t�B�V�X�Ƃقړ�������ł���B
�v���g�A���B�X�͈ȉ��̂悤�ȓ����������Ă���B
�@�@�@�@�E�����̍\�������ɋ߂��B
�@�@�@�@�E��������
�@�@�@�@�E�{�̉��̎��������Ă���i����͎n�c���������ɋ߂��j
�@�@�@�@�E�H�������߂ɋ��̍������B���Ă���i����͎n�c���������ɋ߂��j
�@�@�@�@�E�H�������ߑ傫�ȍ��������B���Ă���B�i����͎n�c���������ɋ߂��j
�@�@�@�@�E��̎w��4�{�i�����������Ɍ��n�I�j
�����̓�������v���g�A���B�X�͎n�c�������������ނɋ߂��̂ł͂ƍl�����Ă���B�v���g�A�r�X�̉����O��I�̒n�w�ɔ����I�̉��������������̂ƍl����l������B�������A�b�|�ނȂǎO��I���L�̐����������ꏊ�Ō������Ă���A�����I�̉������������ƌ����͍̂l���ɂ����B
�����A�v���g�A�r�X�����������n��ł̓_�`���E�����Ƃ͗މ��W�͂Ȃ����_�`���E�����ɂ�������ȃV���{�T�E���X���������Ă���A�܂�Ŕ����I�̐�������肵���������̂悤�ȏꏊ�ł������炵���B
�y�����̉����`���̂P�`�FBCF�iBird�@Came�@First�j���_�z
�@�@�@�@�@�n�c������̎��㔒���I�ɑ��X�o������H�ы����E���`����
�@�@�@�@�A�n�c���ȑO�̎O��I�́h���h�v���g�A�r�X
���̖����������ɉ������邩�B���̉ƂȂ鉼���̈��BCF���_�ł���B�A�����J�̋��������ƃI���V�F�t�X�L�[�ɂ�邱�̗��_�͒P���ɂ����ƁA�R�y���j�N�X�I�Ȕ��z�̓]���ł���B
�܂�A
�@�@�@�@������������i�������̂ł͂Ȃ��A������������i������
�Ƃ������̂ł���B���̗��_�ł͒�����̑S���Ԃ�ʂ��Ď��㐫�̃_�C�m�o�[�h�Ƃ������Ƌ����̋��ʂ̑c�擮����z�肷��B��̃o�[�h�͌������ނւ̐i�����w���������ŁA�ꕔ�̌n���͒n��ɍ~��ċ����ɐi�������Ƃ������ł����B���̗��_�ł͏�̖������ȉ��̂悤�ɉ�������B
�@�n�c���Ȍ�̔����I�ɒ��Ɏ����������o������̂͂Ȃ����B
�����I�̒��`�����͌������ނɂ��߂Â����_�C�m�o�[�h����i���������߁A���Ɏ����`�����p���ł��邩��ɉ߂��Ȃ��B
�A�n�c���ȑO�̎O��I�́h���h�v���g�A�r�X�ɂ��Ăǂ��������邩�H
���łɒ��ւ̐i���̊�{�͎O��I�ɂ͏I����Ă����B�ނ���n�c���̂ق����Â��_�C�m�o�[�h�̌`�����p���������ƍl������B
�������A�����܂œǂ�ł���ꂽ���͂��C�Â��ł��낤�B���̐��ł͐�قnj����e�B���m�T�E���X�̌��߂�����ł��Ȃ����Ƃ��B�����狰�����i�����A���łɎO��I�ɒ��̌��^���o���オ���Ă����Ƃ���A���̃e�B���m�T�E���X�̌��߂͒��̂悤�ɑ傫�������Ȃ��̂ł��낤���B
�����̐i���͈�x�A�������`���͓�x�Ɩ߂�Ȃ��Ƃ��������������Ă���B
�e�B���m�T�E���X�̌��͂��̖@���ɖ������Ă��Ȃ����낤���H�܂�A���������Ă��Ȃ����n�I�Ȍ`���������ŕ������邱�Ƃ͍l���ɂ����B�܂��������_�C�m�o�[�h�̉����������Ă��Ȃ��̂����̐��̏d��Ȍ��_�ł���B
�y�����̉����`���̂Q�`�F�����r�b�N�E�o�����_�z
������x�A���̕\�����Ăق����B
| �O��I | �W�����I | �@�����I | ||
| ��� | �O�� | ��� | �O�� | ��� |
| �w�����T�E���X �R�G���t�B�V�X |
�f�B���t�H�T�E���X �i�P���g�T�E���X�ށj |
�P���g�T�E���X �i�P���g�T�E���X�ށj �A���T�E���X �I���j�g���X�e�X �n�c�� |
�J���m�^�E���X �o���I�j�N�X �H�ы��� �f�B�m�j�N�X �E�q�� |
�K���~���X �e�B���m�T�E���X �e���W�m�T�E���X �I�r���v�g�� �g���G�h�� |
�W�����I�O���̉��L�^���R�������Ƃɂ��C�Â����낤���H���́A�O��I����̑��ł̌�A���������W���[�����Ƃ��Ă̒n�ʂ��m������A�W�����I�O���̉��L�^�͑��̎���ɔ�זR�����̂ł���B
�����r�b�N�o�����_�Ƃ�
�O��I����̑��Ō�`�W�����I�O���ɂ����ċ����͐��Ԍn�ɊJ�������߂邽�߃J���u���A�����I�ȓK�����U�𐋂��Ă����B���̉ߒ��Œ��ɂ����邷�ׂĂ̌n���̋��������łɏo�����Ă����B�����A���̎���̉��L�^���R�������߂ɂ��̂��Ƃ�������Ȃ����������ł���B
�ƌ������̂ł���B
�܂�A�O��I�`�W�����I�O���ɋ����̎傾�����O���[�v�͂��ׂďo�����A�W�����I�̊��ɓK�������푰�i�P���g�T�E���X�ނ�e���k�^�ށA���r�ށA�����ށj�����^�����n�߂��ƌ������̂ł���B�W�����I�O���ɂ́A�b�r�ނł̓_�`���E�����A�e�B���m�T�E���X�ށA�}�j���v�g���ނ̂����Ƃ������̂��̂͂��łɑ��݂������̂́A��^��������Ɍb�܂ꂸ�A���Ԍn�̕Ћ��Ō̐������Ȃ��A�Ђ�����Ə��^�����Ƃ��Đ����Ă����A�����Ĕ����I�ɓ����đ�^���ł�����Ɍb�܂�A�����I�ɔɉh����������������Ȃ��B����́A�n�c������̎��㔒���I�ɑ��X�o������H�ы����E���`�������o������̂́A�����܂��������Ă��Ȃ������ŁA�P�Ȃ���o�ɉ߂��Ȃ��ƌ������Ƃł���B
�����܂ł��ƒP�Ȃ鐄���ɉ߂��Ȃ��B�������A�T�Ƃ��Ĕ����I����̋����̂̑c�悪�W�����I�ɑ��݂����Ⴊ�o�Ă��Ă���B���Ƃ��Ĕ����I����ɔɉh�����A���L���T�E���X�ނ����ނ�̑c�拰���Ǝv����e���`�m�T�E���X�������Ŕ�������Ă���B�܂��A�e�B���m�T�E���X�ނ��W�����I�̃|���g�K���̒n�w����A���B�A�e�B���j�X�E�W�����V�J�Ƃ����c�拰������������Ă���B
���L�^�������ƏW�߂�Α��̌n���ł������悤�ɋN�����n�c���ȑO�ɂ����̂ڂ邱�Ƃ��ł��邩������Ȃ��B
���̉�������{�I�ɂ͐��������A
�@�@�E������������Ă��Ȃ����z�������������Ԃ��W�����I�O�������Ɍ���ł���_
�@�@�E�e�O���[�v�̐i���̃X�e�b�v�Ǝc���ꂽ���n�I�`���̐����ɖ��������Ȃ�����
�Ȃǂ���BFC���_�����L�͂ƍl���邪���������낤���B
�v���g�A�r�X�i�V���{�T�E���X�����������j�ɂ��ẮA���̂悤�Ȑ������ł���̂ł͂Ȃ����낤���B�ނ�͋����������I�Ɏ�ނ��g�債�Ă������ŏo�������A����������s�҂ł͂Ȃ��������낤���B�ނ�͔����I�̒��⋰�������`���������Ȃ�����A�O��I�̊��͂�������ꂸ�A�q�����c�����ƂȂ���ł��Ă��܂����A�k�Ԃł������ƁB
![]()
 �@
�@
�i�f�B�m�j�N�X�S�g�j�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�i�����A�b�v�j

1964�N�A�����I�����s�b�N�̔N�ɔ������ꋰ�����l�b�T���X�̉Εt�����ɂȂ��������ł���B�����Ȋw�����قł͔����҂̃I�X�g�������m�̉����������B
�������A�W�c�Ƃő��H�����̉����ꏏ�ɔ������ꂽ���߁A��둫����܂�ɁA�Q��Ŏ�������ƍl�����Ă���B�m���ɁA����������J���X���炢�̂��Ƃ͂ł������ȋC������B
�������A�܂Ɋւ��Č����Ύ��㐶���҂������c��̖��c�Ƃ�����������B�ĊO�A��Q��ŏ��������F���L���v�g���Ɠ����悤�ɁA�M���ނ�g�J�Q�Ȃǂ̏������𒆐S�Ƃ����H�����ŁA���܂ɑ��̓��H�����̐H�c���⎀�̂������Ă����̂����m��Ȃ��B
�W�c�Ŏ������ɂ��Ă��A�Ԃ蓢���ɂ���悤�ȑ����I�ԂƂ͍l���ɂ����B�ӊO�ƃI�X�g�������m�����������͎̂��̂������Ă��邤���ɁA�˔��I�Ȏ��̂Ɋ������܂�Ă��܂����Q��Ȃ̂�������Ȃ��B
�i�ő�̕���ƌ�����r����܁A���㐶���̖��c�ƌ��������j
�P�D�C���f�b�N�X�ɖ߂�
�Q�D��6��F�����Ȋw�����ف@�V�ف`�����̐i���Ɛ�Ł`
�R�D��8��F�����Ȋw�����ف`�V�فFT�EREX�Ɣ����I�̃v���f�^�[�����`
���ӌ��A�����z��![]() �܂�
�܂�
{kind=link}