�Q���S���E���h�L���ɂ��āi2010-Dec-30�j
���h�L�Ƃ��_�}�V�Ɩ��̂��������́A���������܂��B�����Ă��܂������ɂƂ��ẮA���G�ȋC�����Ǝv���܂��B�Ⴆ�A�T�\�����h�L�̓T�\���̒��ԂłȂ��A�N���ɋ߂���ł��B�q���E�����g�J�Q���h�L���g�J�Q�ł͂Ȃ��A�������̒��Ԃł��B�Ƃ��낪�A�Q���S���E���h�L�́A������Ƃ����Q���S���E�̒��Ԃł��B�������Ȃ���A �Q���S���E���h�L���ƃQ���S���E���i�I�I�Q���S���E��N���Q���S���E�Ȃǁj�Ƃł́A���ފw�I�A���Ԋw�I�ɂ��قȂ�܂��B�Q���S���E��������n�R���ɑ��āA�Q���S���E���h�L���́A�k���n�R���̍����ł��B�����ŕ��z�̋N�����قȂ�܂��B
�Q���S���E���h�L���ƃQ���S���E���i�I�I�Q���S���E��N���Q���S���E�Ȃǁj�Ƃł́A���ފw�I�A���Ԋw�I�ɂ��قȂ�܂��B�Q���S���E��������n�R���ɑ��āA�Q���S���E���h�L���́A�k���n�R���̍����ł��B�����ŕ��z�̋N�����قȂ�܂��B
�Â�������ǂނƁA�Q���S���E���h�L�̎����u�Q���S���E�_�}�V�v�Ƃ����L�ڂɂȂ��Ă�����̂������ł��B���炭�A�����l���Q���S���E�ƌ��ԈႦ��قǎ��Ă��邩�炾�Ǝv���܂��B���������Ă��鎄�ł����A�ȑO�́A�I�I�Q���S���E�A�V���[�v�Q���S���E���h�L�A�K���V�̋�ʂ���o���܂���ł����B�i�O�O�G
���N���ɂ��̃z�[���y�[�W���Љ��ۂɂ́A�u�Q���S���E�_�}�V�̐��E�v�ƌ����A�ʂ��邩������܂���ˁB���̃z�[���y�[�W�̃^�C�g���́A�uDytiscus World�v�ł����A���{��ł́A�u�Q���S���E���h�L���̐��E�v�ƂȂ�܂��B���Ƃ��ẮA�u�Q���S���E���̐��E�v�Ƃ����v�������߂Ẵl�[�~���O�ł��B�܂�A�Q���S���E�ނɍ�������Đڂ��Ă���ƌ����Ӗ����������߂Ă��܂��i�����߂��ł��ˁj�B
���݁A�n����ɂ�26��ނ̃Q���S���E���h�L���iDytiscus�j������Ă���܂��iRoughley, 1990�j�B�啔�����k�����̖k���n��Ɍ��肳��Ă���A�k����Ƃ��Ēm���Ă��܂��B�������Ȃ���A�ԓ������̃��L�V�R��A�����J�̃t�����_�ƌ������悤�Ȓg�����n��ɂ��������Ă���A�Q���S���E���h�L���̐i���n����R�����A�傫�ȃJ�M�������Ă���̂ł͂Ȃ����Ǝv���Ă��܂��B�Q���S���E���h�L���̌`�ԓI�����́A�����i�z�j�ɐԂ�V���̖�l������܂��B���̃Q���S���E�ނƂ̌���I�ȈႢ�ł��B
��ʓI�Ȑ��Ԃ́A�Q���S���E���Ƃقړ��l�ɁA�t����Ăɂ����ĔɐB���A�H���ɐ����ƂȂ�܂��B�������A�V���[�v�Q���S���E���h�L�Ɋւ��ẮA��q�̒ʂ�A���قȐ����j�������Ă��܂��B�܂��A�O���ɂ́A�c�����ɉz�~����B��̎�i�V���[�v�Q���S���E���h�L�̎o����ł���D. semisulcatus�j�����܂��B����ȊO�́A�ʏ�A�Y�������N�ɐ����ɂȂ�悤�Ȑ����j�i�ʏ́A��N�ꉻ�j��L���Ă��܂��B
�H���Ɋւ��ẮA�c�����ɂ͗����ނ̗c���i�I�^�}�W���N�V�Ȃǁj����H�Ƃ��Ă���Ƃ̕������ł����A��ɂ���ẮA�I�^�}�W���N�V�̖L�x�Ȏ��牺�ł��A�쎀���Ă��܂��킪���Ȃ��Ƃ�4��m���Ă���܂��iD. harrisii, D. semisulcatus, D. latissimus, D. circumcintus�j�B�����́A�~�m���V�̂悤�ȏ�ɓ����Ă���Caddis larvae�i�J���Q���ނ̗c���j�@�������ς�ߐH���Ă���܂��B���R�̎��Ȃ���A��A�����n�̊��ɂ���āA�H���A���Ԃ��傫���قȂ�܂��B�����ɂȂ�ƃQ���S���E�Ɠ��l�A�����������A�����ނ�ߑ������ȂǁA��p�ɑO���Œ͂�ŕߐH���܂��B�����ɐ�������Q���S���E���h�L���́A��L4��̂悤�ȋ����n�D���͎����Ă��炸�A��r�I�A���ł��ǂ��H�ׂ���ł��B
���āA���{�����̃Q���S���E���h�L���́A���݁A3��1����m���Ă���܂����i�Q���S���E���h�L�iD. dauricus�j�A�G�]�Q���S���E���h�L�iD. marginalis czerskii�j�A�V���[�v�Q���S���E���h�L�iD. sharpi�j�j�A����̕��q��`�w�̐i���ɔ����A�����A�����Ƒ����̎�ɕ��ނ����\�����ɂ߂č����ł��B
�V���[�v�Q���S���E���h�L�ȊO�́A���ׂāA���k�k������k�C���ɕ��z���Ă���܂��B�V���[�v�Q���S���E���h�L�́A������ł���A�Y�}�Q���S���E���h�L�iD. sharpi�j�ƕʈ���ł���i�R�Q���S���E���h�L�j�ɕ������Ă��܂��B������Ԃł̈Ⴂ�́A���X�̏����ɐ[���āA�����c�a������̂��A�R�Q���S���E���h�L�ł��B�A�Y�}�Q���S���E���h�L�̃��X�ł́A�I�X�Ɠ��l�A���̏c�a���S�������^�C�v���A�����Ă��A�Z���ł��B�I�X�Ɋւ��ẮA�قƂ�ǔ��ʕs�\�ł��B�G�]�Q���S���E���h�L��Q���S���E���h�L�́A���݂ł���r�I�A�̐��͑����ł����A�V���[�v�Q���S���E���h�L�A���ɃA�Y�}�Q���S���E���h�L�Ɋւ��ẮA��Ő��O�̏�ԂƎv���܂��B1931�N�̕����ɂ́A�u�V���[�v�Q���S���E�_�}�V�A�����ɎY����NjH�Ȃ�A�R�Q���S���E�_�}�V�A���s�n���ɎY����ǂ��H�Ȃ�v�B�����ɂ��Â߂��������͂ł����A�V���[�v�Q���S���E���h�L�̕��z�́A���̎��ォ��m���Ă���A����ɋ����[�����Ƃ́A�����ł������A��Ƃ��ċL�ڂ���Ă������Ƃł��B���{�̍��x�������ȍ~�̊������̒���������ł́A�{�킪�����ł�����́A�ʂ����Ăǂ̒��x�c����Ă���̂��Ƒz�����Ă��܂��܂��B
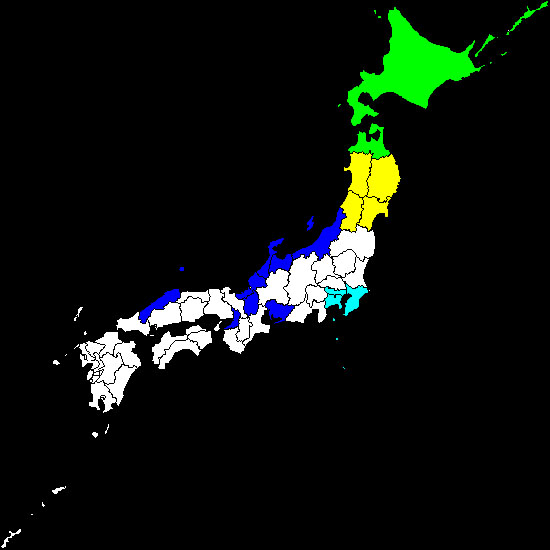 |
| �Q���S���E���h�L���̍������z�i���ɐ�ł����n����܂ށj |
���F�Q���S���E���h�L�A�G�]�Q���S���E���h�L
���F�G�]�i�L�^�j�Q���S���E���h�L
���F�R�Q���S���E���h�L
���F�A�Y�}�Q���S���E���h�L |
�����n��ɐ������Ă��铯����̒��ŁA�V���[�v�Q���S���E���h�L�͊֓��i��t���j��k���i�ΐ쌧�Ȃǁj�̂悤�ȉ��g�Ȓn��ɐ������Ă��܂��B���̓����I�ȕ��z�̓�Ɋւ��āA���̈�A�̌����̒��ňꕔ�𖾂���܂����B���Ȃ킿�A�{��͐����j�̈ꕔ�i�����珉�ߗc�������j�ɉ��x���̍��������������Ă���A���������邽�߂ɑ����̌���A�Y���A�z�����邱�Ƃʼn��g�Ȓn��ɂ��܂��K�����Ă��܂��iInoda, 2003�j�B
�܂��A�{��̐��B�x�������͉��x�ɋ����ˑ����A20���ȉ��ɂȂ�ƌ���s����U������Ɠ����ɃI�X�̐��B���i�����ƕ����B�j���тɗ��������n���A12���܂łɐ��n���������܂��B����Ƀ��X�́A1-2����8���ȉ��̒ቷ�ɖ\�I����邱�Ƃŏ��߂ė��זE�����n���܂��B�܂�A�������̒i�K�I�ȉ��x�ω��i�����I�ɂ́A���Ȃ��Ƃ�2��ށj���V���[�v�Q���S���E���h�L�̐����y�єɐB�ɂ́A�K�v�s���ȗv���ł��邱�Ƃ����炩�ƂȂ�܂����iInoda et al., 2007�j�B����͂������̂́A�Y�����Ȃ������o�����������̃u���[�_�[�͑����Ǝv���܂��B����͂܂��ɁA�i�K�I�Ȑ������߂̃^�C�~���O�Ɏ��s�����T�^��Ƃ�����ł��傤�B
200�V�N�ɔ��\���������̌�������A�V���[�v�Q���S���E���h�L�́A�����̐��B�x�������@�\�����āA�����b���Ƃ��Ă͉��x�����̗v���ł��邱�Ƃ����E�ōŏ��ɔ������ꂽ�����ł��B�܂��A���x�����B�x�������v���ł����ɂ����āA���Y�ł���臒l���قȂ邱�Ƃ��A�V���[�v�Q���S���E���h�L�ŏ��߂Č��o���ꂽ�ɂ߂ă��j�[�N�ȍ����ł��B�Q���S���E���h�L���́u�����ł��邱�Ƃ��w��I�ɋM�d�ȍ����v�ƌ����L�q�͗ǂ��������܂����A���B�x���ƌ��������̐����w�I�ȑ��ʂɂ����Ă��A�w��I�ȏd�v�������邱�Ƃ����炩�ƂȂ�܂����B
�V���[�v�Q���S���E���h�L�̕ߐH�s���Ɋւ��ẮA��ʓI�ɍs��ꂽ�����͂���܂łقƂ�ǂȂ����߁A�c�����̐����n�ɂ����鐶������ʓI���ԗ��I�ɒ��ׁA�a����𐄒肵�Ă݂܂����B���̌��ʁA���}�A�J�K�G���̃I�^�}�W���N�V�ƃ~�Y���V����тʂ��đ����A���ɁA���}�A�J�K�G���̃I�^�}�W���N�V�������n�̗c���o�������i3-4���j�ɂ�����D���ł��邱�Ƃ��킩��܂����iInoda et al., 2009�j�B
�����ŁA���}�A�J�K�G���c�����{��c���̉a��������̂ЂƂ��ƍl���A���牺�ɂ����āA���̃|�e���V�����������ۂ���m�邽�߂ɁA�l�X�Ȏ������s���܂����B���̌��ʁA�@�c���̃T�C�Y�ɉ������œK�ȉa�T�C�Y�����݂��邱�ƁA�A1���̗c���������ɂȂ�܂łɗv�����I�^�}�W���N�V�̐��́A��300�C�A�B���}�A�J�K�G���I�^�}�W���N�V�������a�ɂ��Ď��炵���ۂ̐����T�C�Y�́A���C���h�̂Ɠ��v�w�I�ɍ����Ȃ������B�ȏ�̂��Ƃ���A�{��c�����̉a�́A���}�A�J�K�G���I�^�}�W���N�V�ł��邱�Ƃ�������������܂����B�܂��A���}�A�J�K�G���I�^�}�W���N�V�Ɏ����ő��������~�Y���V�������a�ɂ��Ď��炵���ꍇ�A1-2�߂܂ł͈炿�܂������A3�ߊ��Ԓ��ɑS�ł��܂����B�~�Y���V�����ł͉a�Ƃ��ĕs�\�����Ǝv���܂��B����͋��炭�A3�ߗc���͑�H���̂��߁A1cm���炸�̃~�Y���V�����ł͐����ɕK�v�ȗ{���������I�ɓ��邱�Ƃ��ł��Ȃ������ƍl���Ă���܂��B���p��Ō����ƁA�œK�ۉa���_�ɂ��A�ߐH�s���̃R�X�g�ɔ�ׂāA���邱�Ƃ̂ł���G�l���M�[�����̃x�l�t�B�b�g�Ƃ̃o�����X���ނ荇��Ȃ��ƍl���Ă���܂��B�~�Y���V�̂悤�ȏ����ȉa���a�Ƃ���ꍇ�A�������̉a��ߐH���Ȃ���Ȃ�܂���B���̂��߂ɂ́A�����̈�A�̕ߐH�s���i�T���A�ߊl�A�ېH���j���s���K�v������܂��B���̂��߂ɂ́A����Ɍ����������̃G�l���M�[��K�v������܂����A�~�Y���V���a�ɂ����ꍇ�ɂ́A���̃R�X�g�̕��������̂ł͂Ȃ����Ǝv���܂��B
�]���āA���R���ł��A�{��c���̓��}�A�J�K�G������v�ȉa�����ł���Ǝv���܂��iInoda et al., 2009�j�B�c�O�Ȃ���A�{�킪�������Ă����t���ł́A�{��Ɠ��l�Ƀ��}�A�J�K�G�����d�v�ی쐶���Ɏw�肳��Ă��邽�߁A�a�������̂���łɕm���Ă���ł��B�V���[�v�Q���S���E���h�L���܂߂���́A�쐶���瓾�邱�Ƃ͍���Ȃ��߁A���牺�ő�ʎ�����@���m�����A�����ɋ�������A���牺�Ő��Ԋw�A�����w�A�s���w�A��`�w�A�n�����ފw�����w�I���Ȃ��Ƃ𖾂炩�ɂ��A�����̕ۑS�ɖ𗧂Ă邱�Ƃ��d�v�ł͂Ȃ����ƍl���Ă���܂��B
�V���[�v�Q���S���E���h�L�́A�Q���S���E���iCybister�j�Ƃ͈قȂ�A���n�ɐ������Ă���܂��B�ł͈�̖{��͂ǂ��ɎY������̂ł��傤���H
D. validus�i�R�Q���S���E���h�L�j�́A�K�}�̗t�ɎY������Ƃ͂̕���܂����AD. sharpi�i�V���[�v�Q���S���E���h�L�j�͂ǂ��Ȃ̂ł��傤���H
����10�N�̊ԁA�V���[�v�Q���S���E���h�L�̔ɐB���@�͒��������サ�A�������I�ȃV�X�e����p���邱�ƂŔ�r�I�e�ՂɔɐB���邱�Ƃ��ł���悤�ɂȂ�܂����iInoda and Kamimura, 2004�j�B�܂��A���݂ł́A�ꕔ�̃u���[�_�[�ɂ����ẮA�{��̓�������������I�ȃV�X�e����p���Ă���Ɋȕւȕ��@�ő�ʂ̗c��������s���Ă���܂��iInoda,
unpublished�j�B����������������s���ȑO�̖��Ƃ��āA�����ɂ�������̌��C�ȗ��邩�����̔N�̔ɐB���ʂɒ��ڂȂ��邱�ƂɂȂ�܂��B�R�Q���S���E���h�L�ɂ����ẮA��O�ŃZ���ɎY�����Ă��邱�Ƃ��m�F�������߁A���牺�ɂ����ăZ�����Y���A���Ƃ��Ď������Ƃ���A�Y�����A���픭���������Ƃ���A�Z�����{��̏d�v�ȎY���A���ł͂Ȃ����ƕ��܂����B�i���c�E�s�z�A1999�j�B�C���^�[�l�b�g���ł̓V���[�v�Q���S���E���h�L�̎Y���A���ɃZ�����g���̂���ʓI�ɒm���Ă���܂����A���͉�X���ŏ��Ɍ��o�������Ƃł����B�������Ȃ���A�����n�̎Y�������ɂ̓Z���ȊO�̐������m���Ă��邵�A�Z���ȊO�̐A���ɂ��Y�����邱�Ƃ�����Ă��܂����B�悭�������鐶�Ԋw�̌�����Ƃ��āA�����ɎY������A������ߐH����Ȃǂ̋L�ڂ�_�����ɐ��������邱�Ƃ��ł��܂��B�������Ȃ���A�{���ɂ���ŗǂ��̂ł��傤���H
�{���I�Ȗ��Ƃ��āA�Y������ړI�͂�������̎q�����㐢�Ɏc�����Ƃł��B�u�����ɎY������v�����łȂ��A�Y�����邱�ƂƓ����ɛz�����������Ȃ����Ƃɂ́A��������̎q�����c�����Ƃ��ł��܂���B�������A�Y�����a�킾���̈ꗗ�\����邱�Ƃ���Ϗd�v�Ȃ��Ƃł����A�����͕K�������œK�ȏꏊ�ɎY������Ƃ͌���܂���B���Ɋ�ɂ����ẮA�œK�Ȋ����Ȃ��Ȃ������ʁA��ނȂ��Y�����Ă���\�����傢�ɂ���܂��B�̐�����������O�̊���m�邱�Ƃ��d�v�ł����A���ƂȂ��Ă͎��x���ł��B���l�ɁA�a�����ɂ��Ă��ߐH���Ă��邩��a�����ƒf�肷��̂͂������Șb�ł��B���ɐ������ɂ���c���ɂ����ẮA���h�Ȑ����ɂȂ邱�Ƃ��K�v�ł��B�H�ׂĂ�����Ɛ������邱�Ƃ��m�F���ď��߂ĉa�����Ƃ��Ẵ|�e���V���������Ǝ�������܂��B���Ɏ��牺�ɂ����āA�S���h�{�ɂȂ�Ȃ��a��ߐH���邱�Ƃ�����܂��B�T�C�G���X�Ƃ́A�ꗗ�\���������̂ł͂Ȃ��A������O�̂��Ƃł����A���̖{���i�Y���A����a�̈Ӌ`�j�m�Ɍ��ɂ߂邱�Ƃ��d�v�ł��B
�E�E�E�E�E�@�b���{�����炻��Ă��܂��܂������A�{��ɖ߂��܂��B
���͒��N�̔ɐB�o������A�V���[�v�Q���S���E���h�L�̎Y���ɂ́A�Z���łȂ�������Ȃ����Ƃ��o���I�ɒm���Ă���܂����̂ŁA�Z���̎Y���L�Ӑ����Ȋw�I�ɏؖ����悤�Ǝv���A�ȉ��̎������s���܂����B
3�N�̊ԁA��t���̔ɐB�n�A2�ӏ����ڍׂɒ����������ʁA�{��̎Y�������i3-4���j��8��ނ̐����肷�邱�Ƃ��ł��܂����iInoda, 2011�j�B���̎����͐������Ⴍ�A�A�����ɖR�����ł����iInoda et al., 2007�j�A�t�̎����Ƃ��ėL���ȃZ�������|�I�ɑ����A�D���ł��邱�Ƃ��킩��܂����B�����ŁA���牺�Ő����n�Ɠ��l�ȐA�������Č����A�Y�����ƛz�����ɂ��ďڍׂɌ��������Ƃ���A�ʔ������ƂɃZ���ւ̎Y���n�D�����L�ӂɍ������Ƃ����o���܂����iInoda, 2011�j�B����ɖʔ��������Ƃ��āA�����n�Ŋm�F����8��ނ̐�����P�ƂŎY���A���Ƃ��ė^���A���������Ƃ���A4��ނ̐����ɂقړ����Y�����邱�Ƃ�������܂����B�Ƃ��낪�A�܂��܂��ʔ������ƂɁA�Z���ɎY�������ꍇ���ł��z�������������Ƃ��킩��܂����B�܂�A�V���[�v�Q���S���E���h�L�̎Y���ɂ́A�Z�����Y���A���Ƃ��ďd�v�ł��邱�Ƃ���������܂����B�Z�������������ł́A�z�����̋ɒ[�Ɉ����w���I���_�J�ɂ��Y�����܂����B�œK�ȎY�������S�����݂��Ȃ����ƂŁA���̎�i��I��������Ȃ����Ƃ��e�Ղɑz���ł��܂��B�{���I�Ȋϓ_���猩���ꍇ�A�w���I���_�J�͖{��̎Y���A���Ƃ͌�����Ǝv���܂��B�ł́A��́A�Ȃ��Z�����{��̎Y���A���Ƃ��čœK�Ȃ̂ł��傤���H
���̂��ƂɊւ��Ă��A���ɖʔ������ʂ邱�Ƃ��ł��A���ɘ_���𓊍e���Ă���܂��B�����ꂱ�̏�ŏЉ�ł���Ǝv���܂��̂ł��y���݂ɁI�I
